The Utility of Blood and Bone Marrow Films and Trephine
Biopsy Sections in the Diagnosis of Parasitic Infections
Clare E. Miller1
and Barbara J. Bain2
1 Honorary
Clinical Research Fellow,
Centre for Haematology, 5th Floor, Commonwealth Building, Hammersmith
Hospital campus of Imperial College London, Hammersmith Hospital, 150
Du Cane road, London W12 0HS, UK.
2 St Mary’s Hospital campus of Imperial College
London, St Mary’s Hospital, Praed Street, London W2 1NY, UK.
Corresponding author: Clare
E. Miller. Honorary Clinical Research Fellow, Centre for Haematology,
5th Floor, Commonwealth Building, Hammersmith Hospital campus of
Imperial College London, Hammersmith Hospital, 150 Du Cane road, London
W12 0HS, UK.
clare.miller@imperial.ac.uk
Published: June 1,
2015
Received: March 29, 2015
Accepted: April 29, 2015
Mediterr J Hematol Infect Dis 2015, 7(1): e2015039, DOI
10.4084/MJHID.2015.039
This article is available on PDF format at:

This is an Open Access article distributed
under the terms of the Creative Commons Attribution License
(http://creativecommons.org/licenses/by/2.0),
which permits unrestricted use, distribution, and reproduction in any
medium, provided the original work is properly cited.
|
|
Abstract
The
laboratory
haematologist has a role
in the
diagnosis of parasitic infections.
Peripheral
blood examination is critical
in
the diagnosis of malaria,
babesiosis,
filariasis and trypanosomiasis. Bone marrow examination is
important in the diagnosis of leishmaniasis and occasionally leads to
the diagnosis of other parasitic infections. The detection of
eosinophilia or iron deficiency anaemia can alert the laboratory
haematologist or physician to the possibility of parasitic infection.
In addition to morphological skills, an adequate clinical history is
important for speedy and accurate diagnosis, particularly in
non-endemic areas.
|
Introduction
Microscopic assessment of blood films and bone marrow samples
plays
a key role in the diagnosis of several parasitic infections. Some
organisms e.g. malaria parasites, babesiae, trypanosomes, leishmaniae
and microfilariae, may be directly visualised in the blood film or
marrow, or associated abnormalities such as thrombocytopenia,
eosinophilia or compensatory bone marrow changes may provide diagnostic
clues. Iron deficiency anaemia can be seen as a result of blood loss
from the gastrointestinal tract with chronic
parasitic
infections of the bowel, or from the urinary bladder with chronic
schistosomiasis. Skill is required to detect and accurately
differentiate organisms, particularly when they are scanty.
Concentration techniques such as buffy coat preparation, centrifugation
and filtration can be used to enhance sensitivity. Serological assays
are available for a number of infections, but these should be used as
an adjunct to microscopy, as none is sensitive or specific enough to be
used on its own to establish a diagnosis.[1]
It is important that the
laboratory is informed if there is clinical suspicion of a parasitic
infection, including details of any relevant travel history, in order
to ensure optimal slide preparation and a high index of suspicion on
examining the slides.
Peripheral Blood Films
Malaria. Examination
of
thick and thin blood films remains the primary method of diagnosis of
malaria in most clinical laboratories. It is recommended as the
diagnostic method of choice where the facilities and expertise are
available, particularly in cases of severe malaria.[2]
Compared with
the rapid diagnostic tests (RDTs) which detect parasite-specific
antigens or enzymes, it has the advantage of allowing species to be
determined and parasites to be quantified and may help identify other
causes of fever. Delays in the diagnosis of malaria often occur due to
the diagnosis not being considered promptly; non-specific laboratory
clues include elevated lactate dehydrogenase, presence of atypical
lymphocytes, elevated aspartate transaminase and thrombocytopenia.[3]
Film
preparation:
A thick film is preferable for detection of parasites and a thin film
for species identification. Although malarial parasites may be detected
in May‒Grünwald‒Giemsa-stained blood films, the specific
parasite
and erythrocyte features are more distinguishable at higher pH with
Leishman or Giemsa staining; a rapid Field stain may also be used.
Blood films should be prepared no longer than three to four hours after
blood collection to minimise the risk of distorted morphology and the
potential appearance of parasite stages not normally occurring in the
blood.[4]
Parasite
and Erythrocyte Morphology:
The distinguishing parasite and erythrocyte features
permitting
identification of the different plasmodium species are well
established (Figures 1‒7,
for morphology of Plasmodium
knowlesi and for comprehensive images of other species
see references 4 and 5). It should be noted that Plasmodium knowlesi,
a parasite only occasionally introduced into Europe, can have some
parasites that resemble P.
falciparum and others that resemble P. malariae.
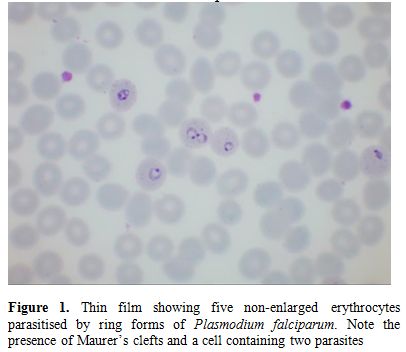 |
Figure
1. Thin film showing five non-enlarged erythrocytes parasitised by ring
forms of Plasmodium falciparum. Note the presence of Maurer’s clefts
and a cell containing two parasites |
|
|
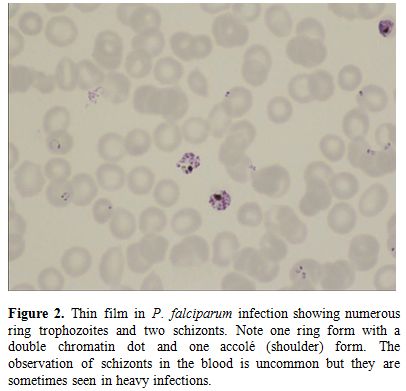
Figure
2. Thin film in P.
falciparum
infection showing numerous ring trophozoites and two schizonts. Note
one ring form with a double chromatin dot and one accolé (shoulder)
form. The observation of schizonts in the blood is uncommon but they
are sometimes seen in heavy infections. |
|
|
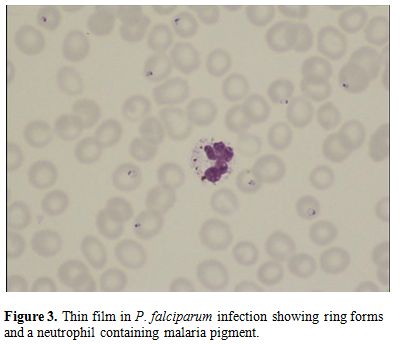
Figure
3. Thin film in P.
falciparum infection showing ring forms and a neutrophil
containing malaria pigment.12:07 01/06/2015 |
|
|
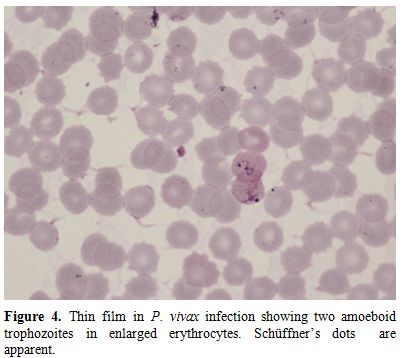
Figure
4. Thin film in P. vivax
infection showing two amoeboid trophozoites in enlarged erythrocytes.
Schüffner’s dots are apparent. |
|
|
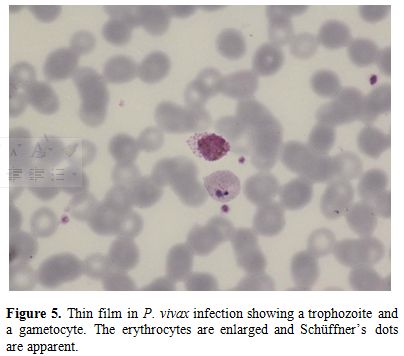
Figure
5. Thin film in P. vivax
infection showing a trophozoite and a gametocyte. The erythrocytes are
enlarged and Schüffner’s dots are apparent. |
|
|
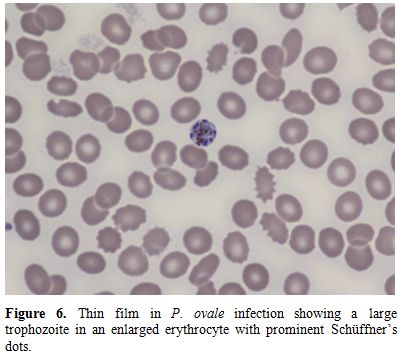
Figure 6. Thin film in P. ovale infection
showing a large trophozoite in an enlarged erythrocyte with prominent
Schüffner’s dots. |
|
|
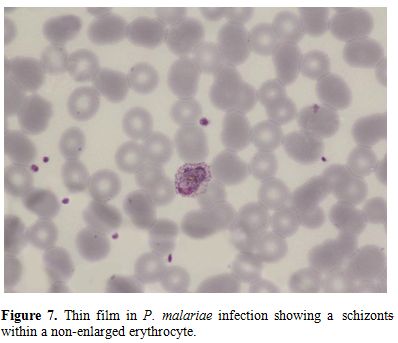
Figure 7. Thin film in P. malariae
infection showing a schizonts within a non-enlarged erythrocyte. |
Accurate laboratory diagnosis of malaria is essential, particularly to
recognise potentially fatal P.
falciparum
infection. Malarial parasites typically appear as cytoplasmic
inclusions within erythrocytes; phagocytosed merozoites and sometimes
schizonts within neutrophils may be seen in P. falciparum
with a high parasitaemia.[6,7]
Parasitized red cells have an altered
appearance, the nature of which varies according to the implicated
species; cells are typically enlarged in P. vivax and P. ovale infections
(Figures 5, 6 and 7).
The malarial pigment, haemozoin, is a degradation product of
haemoglobin and may be seen in monocytes and occasionally
neutrophils (Figure 3).
It can
be visualised readily in stained or unstained films and is
birefringent when polarised light is used.[8]
Monocytes containing
malarial pigment can often be found in the blood for many days after
parasitized red cells have disappeared; this can be useful in making a
retrospective diagnosis of malaria.[9]
In the case of P.
falciparum or P.
knowlesi infection
the degree of parasitaemia should be reported to help assess disease
severity and monitor treatment response. A count of the
proportion
of cells that are parasitized can be made, facilitated by a Miller
graticule, or the number of parasites per ml can be calculated in
relation to the number of white cells. Paradoxically, patients with few
or no parasites detectable on initial blood examination may in fact be
seriously ill due to parasitized red cells being sequestered in
tissues. Parasitaemia is frequently over- or under- estimated and
participation in quality assessment schemes and appropriate referrals
to reference laboratories are important measures to improve
practice.[10,11] All films should
be examined by two people, at least
one of whom should have considerable experience in the field. For
laboratories that do not often see cases of malaria, examination of
films can usefully be supplemented by RDTs.
Associated
abnormalities:
The differential blood count varies considerably between
individuals with malaria. Thrombocytopenia is seen in approximately
60‒80% of people, most commonly but not only in those with P. falciparum or P. knowlesi
infections.[12-14] Possible causes
include reduced platelet survival
from peripheral destruction, enhanced splenic uptake or sequestration,
and decreased platelet production.[15]
Complicating disseminated
intravascular coagulation can occur in falciparum malaria and, rarely,
in vivax malaria.[16] Other
potential findings include a haemolytic
anaemia, leucocytosis or leucopenia, early neutrophilia (with P. falciparum)
or neutropenia, lymphocytosis or lymphopenia (more commonly
lymphopenia) and monocytosis or monocytopenia. Worse prognosis has been
associated with both lymphopenia and lymphocytosis in different
studies. In one study in children a high lymphocyte count and a low
monocyte count were found to correlate with mortality but
thrombocytopenia did not.[7] In a
second study, thrombocytopenia,
leucocytosis and neutrophilia were significantly associated with severe
falciparum malaria in comparison with non-severe and non-falciparum
malaria but the lymphocyte count and the neutrophil: lymphocyte ratio
did not differ between groups; the neutrophil: lymphocyte ratio did,
however, correlate with the degree of parasitaemia.[17]
In a third
study severe malaria was associated with a higher neutrophil:
lymphocyte ratio, a lower lymphocyte count and a lower monocyte count
than non-severe malaria.[18] In
view of the conflicting results in
these and other studies of leucocyte counts, such changes cannot be
regarded as reliable indicators of disease severity. The reticulocyte
count may be inappropriately low as result of bone marrow suppression;
pancytopenia has also been reported.[19]
Clinical and laboratory staff
should also be alert to the possibility of a severe delayed haemolytic
anaemia in patients who are treated with artemisinin.[20]
Atypical
lymphocytes are present in malaria and in some patients with
hyper-reactive malarial splenomegaly.
Babesiosis. Babesiosis is an uncommon tick-borne
parasitic disease caused by a haematoprotozoan of the genus Babesia. Babesia microti
is the commonest causative organism and is endemic in southern New
England, southern New York state, Wisconsin and Minnesota, primarily
occurring between May and October. It is more often detected in
hyposplenic and immunosuppressed patients and the parasitaemia levels
are also usually higher in these patient groups. It is an emerging
threat in transfusion medicine in the United States, with 162 reported
transfusion-associated infections between 1982 and 2013 and 12
associated fatalities in the period 2005‒2008.[21]
B. duncani
has also been transmitted by transfusion.[22]
Transfusion-transmitted
infection, like naturally occurring tick-transmitted infection, is more
often recognised in hyposplenic patients including patients with sickle
cell disease.[23] B. divergens, a
parasite of cattle, causes sporadic cases of babesiosis in the USA,
Europe and Asia, most often in hyposplenic patients. B. bovis infection
also occurs occasionally in Europe.[24]
B. venatorum,
a parasite of roe deer, causes occasional cases in Europe.
Film
preparation: Thick and thin films should be examined as
for malaria.
Parasite
Morphology: The trophozoites of Babesia species are small
rings, easily confused with those of P. falciparum. They
are 1‒5 μm
in diameter with one, two or three chromatin dots and scanty cytoplasm.
Sometimes they are pyriform (pear-shaped) and either paired or have the
pointed ends of four parasites in contact to give a characteristic
Maltese cross formation (see reference 5).
Extracellular parasites may
be seen and can form clusters.[24,25]
B. microti
and B. duncani
trophozoites are indistinguishable morphologically; both are associated
with Maltese cross and ring forms, the latter with small to large
cytoplasmic vacuoles. Their smaller size, vacuolation, polymorphism of
the ring forms, the presence of trophozoites and absence of haemozoin
all help distinguish them from P.
falciparum. Malaria RDTs are negative in babesiosis.
B. divergens and
B. venatorum
typically appear as pyriform pairs of parasites at the periphery of the
erythrocyte but also appear, rarely, as tetrads.[22,26]
Associated
abnormalities:
Babesiosis is often associated with lymphopenia and thrombocytopenia.
Haemolysis is usually mild. There may be atypical lymphocytes.
Trypanosomiasis. African trypanosomiasis (sleeping
sickness) is caused by Trypanosoma
brucei gambiense (West Africa and western Central Africa)
and T. brucei
rhodesiense
(East, Central and Southern Africa). It is transmitted by the tsetse
fly. American trypanosomiasis (Chagas’ disease) is caused by T. cruzi.
Trypanosomes may be detected in the peripheral blood as extracellular
parasites (trypomastigotes). As with malaria, the quality of blood film
microscopy is improved by participation in external quality
assessments.[27]
Film
preparation and staining:
Trypanasomes may be seen moving in a wet preparation when a drop of
anticoagulated blood is placed on a slide, beneath a coverslip, for
microscopic examination. They can also be detected in fixed
preparations such as thick or thin films or buffy coat films. Scanty
parasites are more readily detected by examining the sediment of 10‒20
ml of haemolysed blood. Repeated examinations and concentration
techniques may be needed, particularly for T. brucei gambiense
and T. cruzi.
Preparations should be examined within four hours of sampling. Live
trypanosomes are highly infectious and appropriate laboratory standard
precautions must be adhered to when handling specimens.
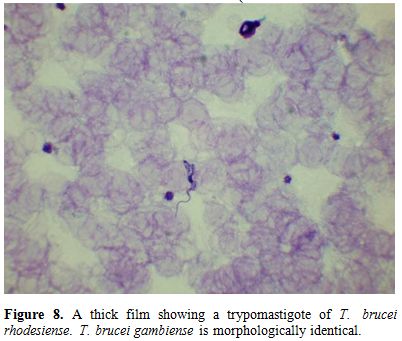
Parasite
morphology: T.
brucei gambiense and T.
brucei rhodesiense
are morphologically indistinguishable, though the latter are more
readily detectable in blood films. They are 13‒42 μm long
with a
slender body, a centrally placed nucleus, a dot- like kinetoplast and a
single flagellum (Figure 8).
The flagellum is joined to the body by an undulating membrane and is
crucial for parasite motility, transmission and pathogenesis.[28] T. cruzi parasites
measure 12‒30 μm and have a larger kinetoplast than the African
trypanosomes. They can be distinguished morphologically from T. rangeli which
has a similar geographical distribution.
Associated
features:
Normocytic normochromic anaemia and thrombocytopenia are often seen
with African trypanosomiasis.[29]
Lymphocytosis and mild anaemia may be
observed in the acute phase of Chagas' disease.
|
|
Figure 8. A thick film showing a
trypomastigote of T.
brucei rhodesiense. T.
brucei gambiense is morphologically identical |
Filariasis. Filariasis
affects over 120 million people worldwide and is endemic in 80
countries.[30] Lymphatic
filariasis is caused by one of three nematodes
(Wuchereria bancrofti,
Brugia malayi and
Brugia timori,
the latter confined to part of Indonesia); filarial infection of the
subcutaneous tissues is caused by Loa Loa. The larvae of these worms,
the microfilariae, are transmitted by mosquitoes to humans, where they
can be found in the blood and show periodicity. W. bancrofti and B. malayi typically
release their microfilariae at night, whereas those of Loa loa are
released during the day.
Film
preparation:
Wet preparations of blood or buffy coat samples may be used for
detection of parasites; examination of a stained film (Giemsa or
another appropriate stain) is needed for determining species.
Concentration methods using centrifugation or stained polycarbonate
filters may enhance detection.[1]
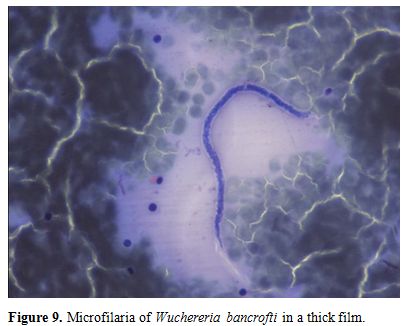
Parasite
morphology:
Microfilariae are classified on the basis of body length and width, the
presence or absence of a sheath, derived from remnants of the egg
membrane, the number of nuclei in the body and the appearance of the
tail including the presence or absence of nuclei in the tail
tip (Figures 9‒11).
In general, pathogenic filariae are sheathed and non-pathogenic are
non-sheathed. However, B.
malayi is sometimes seen unsheathed.[31]
Onchocerca volvulus,
which infects subcutaneous tissues (adult forms) and the eyes
(microfilariae), is occasionally seen in the blood, especially in heavy
infections and after therapy; it is unsheathed with a pointed tail that
lacks nuclei.[31]
Associated
features:
Lymphatic filariasis is typically associated with an eosinophilia;
blood eosinophil count may be used as a nonspecific screening tool in
endemic areas.[32]
|
|
Figure 9. Microfilaria of
Wuchereria bancrofti in a thick film. |
|
|
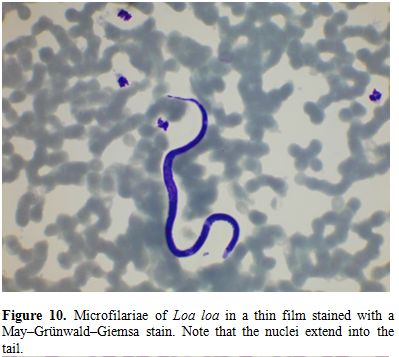
Figure 10. Microfilariae of Loa loa in a thin
film stained with a May‒Grünwald‒Giemsa stain. Note that the nuclei
extend into the tail. |
|
|
Figure 11. Microfilaria of Loa loa in a thin
film stained with Giemsa and Dellafield stain, which shows the sheath
well. |
Others. Rarely, Toxoplasma
gondii has
been identified in the peripheral blood, either extracellularly or
within neutrophils, in patients with toxoplasmosis and underlying
immune deficiency.[33,34]
Phagocytosed leishmaniae (amastigotes) are
occasionally detectable within peripheral blood monocytes or
neutrophils, particularly in immunosuppressed subjects. There may be an
associated pancytopenia, anaemia, leucopenia or thrombocytopenia; red
cell agglutination, fragmentation and rouleaux are also seen.[35]
Bone Marrow Cytology
Malaria.
Malarial parasites may be visualised in red cells or neutrophils or
apparently free in a bone marrow aspirate (see reference 36),
although bone marrow aspiration is not a recommended diagnostic method
for suspected malaria. In acute falciparum malaria the bone marrow may
be hypocellular, normocellular or mildly hypercellular. Immature
gametocytes, which are not usually seen in the peripheral blood, may be
detected in the bone marrow.[37,38]
In chronic falciparum malaria there
is hypercellularity with erythroid hyperplasia. Other features include
dyserythropoiesis, giant metamyelocytes and increased eosinophils,
lymphocytes, plasma cells and macrophages, sometimes with
haemophagocytosis.[6,39,40] The bone marrow in P. vivax
malaria is also characterized by dyserythropoiesis, increased
macrophages (some showing haemophagocytosis), increased plasma cells
and sometimes increased eosinophils.[6]
In hyper- reactive malarial
splenomegaly there may be a marked increase in bone marrow
lymphocytes.[41]
Babesiosis.
Haemophagocytosis has been observed in the bone marrow in babesiosis.[42,43]
Leishmaniasis.
Visceral leishmaniasis is a vector- borne protozoan disease associated
with replication of parasites in macrophages; it is transmitted by
female sandflies. Bone marrow aspiration is very useful in the
diagnosis of visceral leishmaniasis and is a recommended diagnostic
method when this is suspected. Leishmaniasis usually results from Leishmania donovani
in the Indian subcontinent, Asia and Africa (in adults and children) or
from L. infantum
in the Mediterranean region and southwest and central Asia; in South
America this same species is known as L. chagasi,
infection being seen primarily in young children and immunosuppressed
individuals. Other species e.g. L.
tropica in the middle east and L. amazonensis
in South America are occasionally viscerotropic; all may be detected by
bone marrow examination.[44,45]
Leishmaniasis is increasingly been seen
in the context of HIV co-infection and generally represents
reactivation of previously subclinical infection.[45]
It is
occasionally seen as a cause of pancytopenia even in patients living
outside areas of endemicity and without a specific travel
history.[46,47]
Diagnostic
sensitivity for splenic, bone marrow and lymph node aspirate smears is
>95%, 55‒97% and 60% respectively.[45,48] Aspirate films can
be stained with a Giemsa, May-Grünwald-Giemsa or Leishman
stain.
Amastigote forms, called LD bodies, may be visualised; they are
characterised by a small paranuclear rod-like body known as the
kinetoplast, giving the organism a characteristic 'double-dot'
appearance (Figure 12).
Leishmaniae are obligatory intracellular parasites of mononuclear
phagocytes, but they may appear more abundant extracellularly due to
disruption of macrophages during spreading of aspirate films.[49]
Increased macrophages, plasmacytosis and erythroid hyperplasia are seen
in the majority of cases.[35]
Dyserythropoiesis can be striking, to the
extent that misdiagnosis as myelodysplastic syndrome has occurred when
the parasites have been overlooked.[50,51]
Plasma cells (including Mott
cells and cells containing crystals or Russell bodies),
dysmyelopoiesis, free floating cytoplasm and intracellular LD bodies in
cells other than histiocytes (polymorphs, metamyelocytes) are uncommon
features.[35,52]
Increased eosinophils and eosinophilic precursors are
seen in 15‒27% of cases.[35,53] There is associated
haemophagocytosis
in up to 75% of individuals; diagnosis can be challenging due
to
overlapping clinical features. Aspirates are often reported as negative
for LD bodies at disease onset, but in our experience LD bodies may be
present but missed because they are infrequent or there was not a high
index of suspicion.[54]
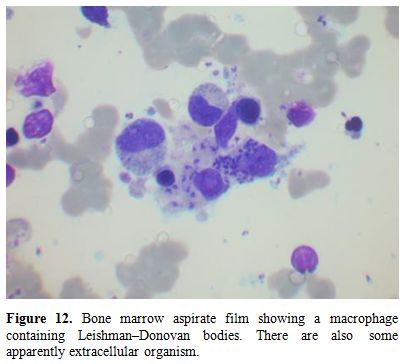 |
Figure12.
Bone marrow aspirate film showing a macrophage containing
Leishman‒Donovan bodies. There are also some apparently extracellular
organisms. |
Others.
Trypanosomes are sometimes detected in the bone marrow, but less often
than leishmaniae. Detection is more common in immunosuppressed
patients.[55,56] Microfilaria are
occasionally observed (see ref
36), also more commonly in the
immunocompromised host; there may be
associated marrow hypoplasia.[57,58]
Toxoplasma have also sometimes
been found in immunodeficient patients, either as free organisms (see
ref 36) or within cysts.[59] An increased number of bone marrow
eosinophils and their precursors are often seen with helminth
infections.
Bone Marrow Histology
A
bone marrow trephine biopsy is not a recommended method for the
diagnosis of parasitic infections but it is necessary to recognise the
histological features since this is occasionally an unexpected
diagnosis, for example when a biopsy is done to investigate fever,
pancytopenia or hepatosplenomegaly.
Malaria.
Bone marrow histology in malaria typically reveals hypercellularity
with increased macrophage activity, often with haemophagocytosis. The
unstained bone marrow films of patients who have had repeated bouts of
malaria may appear slate grey or black because of the accumulation of
haemozoin.[6] It is important to
distinguish haemozoin from formalin
pigment. Haemozoin may be seen not only in macrophages but also within
erythroid and granulocytic precursors, possibly contributing to
dyserythropoiesis and erythroid suppression.[60]
There is correlation
between the amount of haemozoin deposition and the severity of anaemia
in children with P. falciparum infection.[61] During attacks of
acute malaria,
sinusoids may be packed with parasitized red cells.[39]
Gametocytes at
different maturation stages can be identified in haematoxylin and eosin
(H&E)-stained sections, progressing from immature leaf-shaped
forms
to mature forms with a more crescentic shape.[62]
The majority of
immature gametocytes may be observed in extravascular spaces, whilst
most mature gametocytes are typically seen in intravascular
spaces.[62,63]
Leishmaniasis.
The bone marrow is hypercellular in the majority of cases of visceral
leishmaniasis; numerous LD bodies are typically present and allow the
distinction from haematological malignancies which can present with a
similar clinical picture. LD bodies appear as 1‒3 µm round bodies
inside macrophages; their morphological features are often less
apparent than when the parasites are visualised in an aspirate. They
are sometimes confused with the fungus Histoplasma capsulatum in view
of their small size. However, leishmaniae fail to stain with periodic
acid-Schiff (PAS) or silver stains and an H&E or Giemsa stain
will
demonstrate the ‘double-dot’ of the nucleus and kinetoplast. Additional
findings include necrosis, noncaseating granulomas, increased fibrotic
foci and increased vascularity. Estimations of the frequency of these
findings have varied considerably between studies, perhaps reflecting
differences in average parasite densities between regions.[35,52,53,64]
Others.
Toxoplasma infections may be detected in bone marrow trephine
specimens. In immunocompetent individuals the only finding may
be
of granuloma formation. In
immunodeficient patients T. gondii organisms
are occasionally seen. They usually take the form of tachyzoites, which
are 3‒6 μm in diameter and have a tiny single nucleus. Occasionally,
cysts containing numerous bradyzoites
are present. Tachyzoites are
negative with a PAS reaction, whereas cysts
or
bradyzoites are generally well recognised by this staining.[65]
Immunohistochemistry is useful to confirm T. gondii infection
and to discriminate the parasite from cellular debris.[65]
T. cruzi may
be detected in trephine biopsy sections from immunosuppressed patients
with acute Chagas disease.[56]
Very rarely schistosomal eggs have been
observed in a trephine biopsy section.[66]
Pneumocystis jirovecii
can involve the bone marrow, particularly in immunocompromised hosts,
but this organism has now been recognised as a fungus rather than a
protozoan.[67,68]
Conclusion
Peripheral blood examination is critical in the diagnosis of
malaria, babesiosis, filariasis and trypanosomiasis but it is also
important to be aware of the possibility of diagnosis of these
infections from bone marrow aspirates or trephine biopsy sections. In
the case of leishmaniasis, it is bone marrow examination that is of
major diagnostic importance, while organisms are only rarely detected
in the peripheral blood.
References
- Rosenblatt
JE. Laboratory diagnosis
of infections due to blood and tissue parasites. Clin Infect Dis 2009;
49: 1103-1108. http://dx.doi.org/10.1086/605574
PMid:19691431
 .
.
- World
Health Organisation. Guidelines for the treatment of malaria.
Second edition. Available at:
http://whqlibdoc.who.int/publications/2010/9789241547925_eng.pdf?ua=1
(accessed 17 May 2015) .
- Cunha BA.
The diagnosis of imported malaria. Arch Intern Med 2001; 161:
1926-1928. PMid:11493157 .
- Warhurst
DC, Williams JE. ACP Broadsheet no 148. July 1996. Laboratory diagnosis
of malaria. J Clin Path 1996; 49: 533-538. PMid:8813948
PMCid:PMC500564 .
- Bain BJ.
Blood Cells: A Practical Guide. 5th
ed. Oxford: Wiley- Blackwell; 2014. pp 152-163.
- Wickramasinghe
SN, Abdalla SH. Blood and bone marrow changes in malaria. Baillière’s
Best Pract Res Clin Haematol 2000; 13, 277-299. PMid:10942626
.
- Ladhani
S, Lowe B, Cole AO, Kowuondo K, Newton CRJC. Changes in white blood
cells and platelets in children with falciparum malaria: relationship
to disease outcome. Br J Haematol 2002; 119: 839-847.
PMid:12437669 .
- Lawrence
C. Laveran remembered: malaria haemozoin in leucocytes. Lancet 1999;
353: 1852. PMid:10359416 .
- Day
NP, Pham TD, Phan TL et al. Clearance kinetics of parasites and
pigment-containing leukocytes in severe malaria. Blood 1996; 88:
4694-4700. PMid:8977263 .
- Manser M,
Olufsen C, Andrews N, Chiodini PL. Estimating the parasitaemia of
Plasmodium falciparum: experience from a national EQA scheme.
Malaria J 2013; 12: 428.
http://dx.doi.org/10.1186/1475-2875-12-428 PMid:24261625.
PMCid:PMC4222811 .
- Bailey
JW,
Williams J, Bain BJ, Parker-Williams
J,
Chiodini PL; General Haematology Task Force of the British Committee
for Standards in Haematology. Guideline:
the
laboratory diagnosis of malaria. Brit J Haematol 2013; 163,
573-580. http://dx.doi.org/10.1111/bjh.12572
PMid:24219330 .
- Srichaikul
T. Hemostatic alterations in malaria. Southeast Asian J Trop Med Public
Health 1993; 24: Suppl 1, 86-91. PMid:7886615 .
- Mueller
I, Galinski MR, Baird JK et al. Key gaps in the knowledge of Plasmodium
vivax, a neglected human malaria parasite. Lancet Infect Dis 2009; 9:
555‒566.
http://dx.doi.org/10.1016/S1473-3099(09)70177-X PMid:19695492
.
- Seilmaier
M, Hartmann W, Beissner M et al. Severe Plasmodium knowlesi infection
with multi-organ failure imported to Germany from Thailand/Myanmar.
Malaria J 2014; 13: 422.
http://dx.doi.org/10.1186/1475-2875-13-422
PMid:25367021
PMCid:PMC4228098 .
- Angchaisuksiri
P.
Coagulopathy in malaria. Thromb Res 2014; 133: 5-9.
http://dx.doi.org/10.1016/j.thromres.2013.09.030
PMid:24099998 .
- Bain B.
Disseminated intravascular coagulation in benign tertian malaria. Brit
Med J 1973: 1: 550 PMid:4571188 PMCid:PMC1588695 .
- van
Wolfswinkel ME, Vliegenthart-Jongbloed K, de Mendonça Melo M et al.
Predictive value of lymphocytopenia and the neutrophil-lymphocyte count
ratio for severe imported malaria. Malar J 2013; 12; 101.
http://dx.doi.org/10.1186/1475-2875-12-101 PMid:23506136
PMCid:PMC3608093 .
- Berens-Riha
N,
Kroidl I, Schunk M et al. Evidence for significant
influence
of host immunity on changes in differential
blood count during malaria. Malar J 2014; 13: 155.
http://dx.doi.org/10.1186/1475-2875-13-155
PMid:24758172.
PMCid:PMC4021259 .
- Gupta SK,
Jain D, Singh T. Plasmodium falciparum in trephine biopsy. Am J Hematol
2008; 83: 602. PMid:17580333 .
- Jarvis
JN, Coltart CE, Pule M, Chiodini PL, Doherty T. Artemisinin therapy and
severe delayed haemolysis. Lancet 2013; 382: 180.
http://dx.doi.org/10.1016/S0140-6736(13)60812-0
PMid:23849926 .
- Lobo
CA, Cursino-Santos JR, Alhassan A, Rodrigues M. Babesia: An Emerging
Infectious Threat in Transfusion Medicine. PLoS Pathog 2013; 9:
e1003387. http://dx.doi.org/10.1371/journal.ppat.1003387
PMid:23853577
PMCid:PMC3708872 .
- Vannier
E, Krause PJ. Human babesiosis. N Engl J Med 2012; 366, 2397-2407. http://dx.doi.org/10.1056/NEJMra1202018
PMid:22716978 .
- Tonnetti
L, Eder AF, Dy B et al. Transfusion-transmitted Babesia microti
identified through hemovigilance. Transfusion 2009; 49: 2557-2563.
http://dx.doi.org/10.1111/j.1537-2995.2009.02317.x PMid:19624607
.
- Setty
S, Khlalil Z, Schori P, Azar M, Ferrieri P. Babesiosis: two atypical
cases from Minnesota and a review. Am J Clin Path 2003; 120: 554-559.
PMid:14560566 .
- Rajpal
DR, Murray
DR, Morrell DR, O’Dwyer DR. Human babesiosis: an unusual cause of
haemolytic anaemia. Br J Haematol 2005; 129: Suppl. 1, 51. .
- Moody AH,
Chiodini PL. Methods for the detection of blood parasites. Clin Lab
Haematol 2000; 22: 189-202. PMid:11012630 .
- Mukadi
P, Gillet P, Lukuka A et al. External quality assessment of
Giemsa-stained blood film microscopy for the diagnosis of malaria and
sleeping sickness in the Democratic Republic of the Congo. Bull World
Health Organ 2013; 91: 441-448.
http://dx.doi.org/10.2471/BLT.12.112706 PMid:24052681
PMCid:PMC3777151 .
- Langousis
G, Hill
KL. Motility and more: the flagellum of Trypanosoma brucei. Nat Rev
Microbiol. 2014; 12: 505-18. http://dx.doi.org/10.1038/nrmicro3274
PMid:24931043 PMCid:PMC4278896 .
- Moore
AH, Ryan ET, Waldron MA. A 37-year-old man with fever,
hepatosplenomegaly and a cutaneous foot lesion after a trip to Africa.
N Engl J Med 2002; 346: 2069-2077. PMid:12087144. .
- Kaushal
S, Iyer VK, Mathur SR. Morphological variations of Wuchereria bancrofti
in cytology smears: A morphometric study of 32
cases. Acta
Cytol 2012; 56: 431-438. http://dx.doi.org/10.1159/000337446
PMid:22846330 .
- Cheesbrough
M.
Learning bench aid series number 3. Microscopical Diagnosis of
Lymphatic filariasis, Loiasis, Onchocerciasis. In: Tropical Health
Technology. Doddington, Cambridgeshire 2011. .
- Musso
D, Vialette C. Predictive value of the eosinophil counts in the
biological diagnosis of lymphatic filariasis in French Polynesia. Med
Mal Infect 2012; 42: 585-590.
http://dx.doi.org/10.1016/j.medmal.2012.09.006
PMid:23116705 .
- Albrecht
H, Sobottka I, Stellbrink H-J, van Lunzen J, Greten H. Diagnosis of
disseminated toxoplasmosis using a peripheral blood smear. AIDS 1996;
10: 799-800. PMid:8805877 .
- Arnold
SJ, Kinney MC, McCormick MS, Dunmer S, Scott MA. Disseminated
toxoplasmosis: unusual presentations in the immunocompromised host.
Arch Pathol Lab Med 1997; 121: 869- 873.
PMid:9278617 .
- Chandra
H, Chandra S, Kaushik RM. Visceral leishmaniasis with associated
common, uncommon, and atypical morphological features on bone marrow
aspirate cytology in nonendemic region. J Trop Med 2013; 861032
http://dx.doi.org/10.1155/2013/861032.
PMid:24089618 PMCid:PMC3782059 .
- Bain BJ,
Clark DM, Wilkins BS. Bone Marrow Pathology. 4th
ed. Oxford: Wiley-Blackwell; 2010. p. 118-124. .
- Smalley
ME, Abdalla S, Brown J. The distribution of Plasmodium falciparum in
the peripheral blood and bone marrow of Gambian children. Trans R Soc
Trop Med Hyg 1981; 75: 103-105. PMid:7022784 .
- Abdulsalam
AH, Sabeeh N, Bain BJ. Immature Plasmodium falciparum gametocytes in
bone marrow. Am J Hematol 2010; 85: 943.
http://dx.doi.org/doi/10.1002/ajh.21796
PMid:20687103 .
- Phillips
RE, Pavsol G. Anaemia of Plasmodium falciparum malaria. Baillière’s
Clin Haematol 1992; 5: 315–330. PMid:1511178 .
- Abdalla
SH. Hematopoiesis in human malaria. Blood Cells 1990; 16: 401-416.
PMid:2257320 .
- Bates
I, Bedu-Addo G, Bevan DH, Rutherford TR. Use of immunoglobulin gene
rearrangements to show clonal lymphoproliferation in hyper-reactive
malarial splenomegaly. Lancet 1991; 337: 505–507.
PMid:1671888 .
- Slovut
DP, Benedetti E, Matas AJ. Babesiosis and hemophagocytic syndrome in an
asplenic renal transplant recipient. Transplantation 1996; 62: 537-539.
PMid:8781622 .
- Poisnel
E, Ebbo M,
Berda-Haddad Y et al. Babesia microti: an unusual travel-related
disease. BMC Infect Dis 2013; 13: 99.
http://dx.doi.org/doi/10.1186/1471-2334-13-99
PMid:23432953.
PMCid:PMC3598249 .
- Sacks DL,
Kenney RT, Kreutzer RD et al. Indian kala-azar caused by Leishmania
tropica. Lancet 1995; 345: 959–961. PMid:7715298 .
- Murray
HW, Berman JD, Davies CR, Saravia NG. Advances in leishmaniasis. Lancet
2005; 366: 1561-1577. PMid:16257344 .
- Drexler
B, Holbro A. Unexpected bone marrow finding in a patient with
pancytopenia after hematopoietic stem cell transplantation. Blood 2014;
124: 678. PMid:25221805 PMCid:PMC4118483 .
- Bogdan
C, Schönian G, Bañuls AL et al. Visceral leishmaniasis in a
German child who had never entered a known endemic area: case report
and review of the literature. Clin Infect Dis 2001; 32: 302- 306.
PMid:11170923 .
- Srivastava
P, Dayama
A, Mehrotra S, Sundar S. Diagnosis of visceral leishmaniasis. Trans R
Soc Trop Med Hyg 2011; 105, 1–6.
http://dx.doi.org/doi/10.1016/j.trstmh.2010.09.006
PMid:21074233 .
- Chopra
A, Anand M, Kalita D, Singh S, Kumar R. Greater abundance of
extracellular Leishmania donovani bodies: possible clue from comparison
of bone marrow aspirate and imprint findings. J Clin Path 2009; 62:
574‒575. http://dx.doi.org/10.1136/jcp.2009.064709
PMid:19474363 .
- Kopterides
P, Halikias S, Tsavaris N. Visceral leishmaniasis masquerading as
myelodysplasia. Am J Hematol 2003, 74:198– 199. PMid:14587050
.
- Bain BJ.
Dyserythropoiesis in visceral leishmaniasis. Am J Hematol 2010; 85:
781. PMid:20652969 .
- Bhatia
P, Haldar D, Verma N et al. A case series highlighting the relative
frequencies of the common uncommon and atypical/unusual hematological
findings on bone marrow examination in cases of visceral leishmaniasis.
Mediterr J Hematol Infect Dis 2011; 3: e2011035.
http://dx.doi.org/10.4084/MJHID.2011.035
PMid:22084650 PMCid:PMC3212968 .
- Daneshbod
Y, Dehghani SJ, Daneshbod K. Bone marrow aspiration findings
in
kala-azar. Acta Cytol 2010; 54, 12-24. PMid:20306983 .
- Rajagopala
S, Dutta U, Chandra KS et al. Visceral leishmaniasis associated
hemophagocytic lymphohistiocytosis--case report and systematic review.
J Infect 2008; 56: 381-388.
http://dx.doi.org/10.1016/j.jinf.2008.02.013
PMID:18405976 .
- Kirchhoff
LV. American trypanosomiasis (Chagas’ disease) – a tropical disease now
in the United States. N Engl J Med 1993; 329: 639–644.
PMID:8341339 .
- Baena
Terán RB, Arancibia A, Basquiera AL et al. Trypanosoma cruzi in the
bone marrow. Br J Haematol 2012; 157:1.
http://dx.doi.org/10.1111/j.1365-2141.2012.09049.x
PMid:22329435 .
- Molina
MA, Cabezas MT, Gimenez MJ. Mansonella perstans filariasis in a HIV
patient: finding in bone marrow. Haematologica 1999; 84: 861.
PMid:10477464 .
- Sharma S,
Rawat A,
Chowhan A. Microfilariae in bone marrow aspiration smears, correlation
with marrow hypoplasia: a report of six cases. Indian J Pathol
Microbiol 2006; 49: 566-568. PMid:17183857 .
- Soulier-Lauper
M, Zulian G, Pizzolato G et al. Disseminated toxoplasmosis in a
severely immunodeficient patient: demonstration of cysts in bone marrow
smears. Am J Hematol 1991; 38: 324-326. PMid:1746542 .
- Casals-Pascual
C, Kai O, Cheung JO et al. Suppression of erythropoiesis in malarial
anemia is associated with hemozoin in vitro and in vivo. Blood 2006;
108: 2569–2577. PMid:16804108 .
- Aguilar
R, Moraleda C, Achtman AH, et al. Severity of anaemia is associated
with bone marrow haemozoin in children exposed to Plasmodium
falciparum. Brit J Haematol 2014; 64: 877-887.
http://dx.doi.org/10.1111/bjh.12716
PMid:24386973 .
- Farfour
E, Charlotte F, Settegrana C, Miyara M, Buffet P. The extravascular
compartment of the bone marrow: a niche for Plasmodium falciparum
gametocyte maturation? Malar J 2012; 11: 285.
http://dx.doi.org/10.1186/1475-2875-11-285 PMid:22905863
PMCid:PMC3472181 .
- Joice
R, Nilsson SK, Montgomery J et al. Plasmodium falciparum transmission
stages accumulate in the human bone marrow. Sci Transl Med 2014; 6,
244re5. http://dx.doi.org/10.1126/scitranslmed.3008882
PMid:25009232 .
- Kumar
PV, Vasei M, Sadeghipour A et al. Visceral leishmaniasis: bone marrow
biopsy findings. J Pediatr Hematol Oncol 2007; 29: 77–80.
PMid:17279002 .
- Brouland
J-P, Audouin J, Hofman P et al. Bone marrow involvement by disseminated
toxoplasmosis in acquired immunodeficiency syndrome: the value of bone
marrow trephine biopsy and immunohistochemistry for the diagnosis. Hum
Pathol 1996; 27: 302-306. PMid:8600047 .
- Jones
C, Leday TV. Schistosomal eggs identified on bone marrow biopsy. Blood
2014; 124: 1219. http://dx.doi.org/10.1182/blood-2014-06-582767.
PMid:25285348 PMCid:PMC4141513 .
- Grier DD,
Lewis Z, Palavecino EL. Bone marrow involvement by Pneumocystis
jiroveci. Br J Haematol 2009; 145: 149. PMid:19533848 .
- Stringer
JR, Beard CB, Miller RF. Spelling Pneumocystis jirovecii. Emerg Infect
Dis 2009; 15: 506. http://dx.doi.org/10.3201/eid1503.081060
PMid:19239784 PMCid:PMC2681121 .
[TOP]