Emmanouil Spanoudakis1, Menelaos Papoutselis1,Ioanna Bazdiara1, Eleftheria Lamprianidi1, Xrisa Kordella1, Constantinos Tilkeridis2, Costas Tsatalas1 and Ioannis Kotsianidis1.
1 Department of Hematology, Democritus University of Thrace, Alexandroupolis, Greece.
2 Department of Orthopedics, Democritus University of Thrace, Alexandroupolis, Greece.
Correspondence to: Emmanouil
Spanoudakis, Assistant Professor of Hematology. Democritus University
of Thrace, Area of Dragana, Alexandroupolis, Greece, PC: 68100. Tel:
00302551351512. E-mail:
emmanouilspanoudakis@yahoo.com
Published: November 1, 2018
Received: July 3, 2018
Accepted: September 16, 2018
Mediterr J Hematol Infect Dis 2018, 10(1): e2018058 DOI
10.4084/MJHID.2018.058
This is an Open Access article distributed
under the terms of the Creative Commons Attribution License
(https://creativecommons.org/licenses/by-nc/4.0),
which permits unrestricted use, distribution, and reproduction in any
medium, provided the original work is properly cited.
|
|
Abstract
JAK2V617F
is a gain of function point mutation that occurs in Myeloproliferative
Neoplasm (MPN) patients and deranges their hemopoiesis at cellular
level. We speculate that hyperfunctioning JAK2 can modify osteoclast
(OCL) homeostasis in MPN patients. We studied
18 newly diagnosed
MPN patients and four age-matched normal donors (ND). Osteoclast
forming assays started from selected monocytes also and under titrated
concentrations of the JAK2 Inhibitor AG-490 (Tyrphostin). Genomic DNA
was extracted from the formed osteoclasts, and the JAK2V617F/JAK2WT genomic DNA ratio was calculated. OCLs formed from monocytes derived from heterozygous (Het) for the JAK2V617F mutation MPN patients, were three times more compared to those from JAK2 wild type (WT) MPN patients (p=0,05) and from ND as well (p=0,03). The ratio of JAK2V617F/JAK2WT
genomic DNA was increased in OCLs compared to the input monocyte cells
showing a survival advantage of the mutated clone. In comparison to ND
and JAK2 WT MPN patients, OCLs from patients JAK2V617F (Het) were more susceptible to JAK2 inhibition. These alterations in osteoclast homeostasis, attributed to mutated JAK2, can deregulate the hemopoietic stem cell niche in MPN patients.
|
Introduction
Osteoclasts
(OCLs) are bone resorting cells of hemopoietic origin. The balanced
function between osteoclasts and osteoblasts determines bone
homeostasis. Osteoblasts are considered important cells for the
hemopoietic stem cell (HSC) niche formation and regulation,[1]
while osteoclasts are considered dispensable cells in this setting.
Their role is to mobilise hemopoietic progenitor cells towards
the peripheral blood especially under stress conditions.[2-6]
JAK2
is an intracellular kinase, a cellular hub, that acts as a second
mediator of intracellular signaling from a variety of cytokine and
hormone receptors.[7,8] Pharmacological inhibition of the JAK2 kinase reduces osteoclast bone resorting activity,[9-12] suggesting that the intact JAK2 function is essential for normal osteoclast function.
The JAK2V617F
is a gain of function point mutation that often occurs in patients with
myeloproliferative neoplasms (MPN) and deranges basic cellular
functions of their hemopoietic cells.[13,14] JAK2V617F
arises in an early myeloid progenitor cell, cells ancestor to OCLs.
Except intrinsic stem cell defects, HSC niche deregulation in JAK2V617F mutated mice models of MPN contributes equally to MPN disease evolution.[1,15]
Therefore we speculate that genetically modified JAK2,
as it occurs in MPN patients, can slightly modify osteoclast
homeostasis in a way that deregulates hematopoietic stem cell niche
homeostasis. Patient’s Samples and Materials
Approximately,
60 ml of peripheral blood was drawn, after informed contest form was
signed, from 18 newly diagnosed MPN patients (14 essential
thrombocythemia, two primary myelofibrosis, two polycythemia vera
diagnosed according to 2008 WHO criteria) and from 4 age- matched
normal donors (ND). Positive selection of CD14+ monocyte cells was
performed by using immunomagnetic beads separation technique
(MiltenyiBiotec). The selective JAK2 inhibitor, TyrphostinAG-490, was
provided by Sigma Diagnostics and was dissolved in DMSO. This study was
approved by the local ethics committee and conducted according to the
principles of the declaration of the Helsinki.
JAK2V617F mutation detection in our patient’s cohort.
Genomic DNA was extracted from whole blood (QIAGEN), and 25 ng of gDNA
were subjected to Allele-specific PCR (ASO-PCR) for the detection of
the JAK2V617F mutation
according to the protocol described by Baxter et al., slightly
modified. Restriction Length Polymorphism method (RFLP) by using the
BsaXI restriction enzyme, was performed in all positive for the JAK2V617F mutation samples, to determine its presence in heterozygous or homozygous status.
Osteoclast (OCL) forming assays from monocyte cells derived from MPN patients. 2x105
positively selected monocyte cells, after checked for purity
by flow cytometry,
were suspended in a MEM medium + 10% Fetal Calf
Serum, supplemented with optimum concentrations of osteclastogenic
cytokines (RANKL 25 ng/ml and M-CSF 50 ng/ml) and then they were plated
in a 48-flat bottom well plate. The half medium exchange was performed
twice weekly, and the number of formed OCLs was scored under inverted
microscope by using TRAP assays (Sigma Diagnostics) after staying 14
days in culture.
Quantitative Real-Time PCR (RT-PCR) for JAK2V617F/JAK2WT genomic DNA ratio measurement.
Genomic DNA was extracted (QIAGEN) from both the input for OCL forming
assays monocyte cells as well as from the OCLs that they were finally
formed from them. JAK2V617F/JAK2WT
genomic DNA ratio was calculated by using the Taqman based
JAK2MutaQuant Kit (IPSOGEN). Briefly, a pair of FAM-tracked
hybridisation probes was used in two different reaction tubes, one of
them specific for the G1849T JAK2 mutation and the other specific for the wild type sequence, together with a JAK2gDNA specific pair of primers. Appropriate serial dilutions of known numbers of mutated and wild type JAK2
transcripts were used in order to set the calibration plot. All assays
were performed on an ABI Prism7500 analyser (Applied- Biosystems).
OCL forming assays in the presence of the JAK2 inhibitorAG-490(Tyrphostin).
Initially, titration of the inhibitory dose of AG-490 on OCL formation
assays was performed. Selected monocytes from three normal donors
started OCL forming assays in the presence of serial dilutions of
AG-490 dissolved in DMSO; range 0.01-100 μM. The 50% Inhibitory
concentration (IC50) was determined at 20μM (Figure 2A).
Then OCL forming assays were started from MPN patient’s monocytes as
previously described, and concurrently OCL forming assays in the
presence of 2μM and 20μM of the JAK2 inhibitor AG-490 (Tyrphostin)
dissolved in DMSO were also started.
CTXβ in serum measurement by ELISA and Colony Forming Assays from CD34 selected cells from the peripheral blood.
CTXβ levels in whole blood serum were measured by ELISA (β-
crossLaps/serum kit, Roche Diagnostics). Data are expressed as ng/ml.
Statistical analysis.
All values represent median values (min-max). Comparisons between
groups were performed by using the Mann-Whitney for unpaired and the
Wilconxon Rank test for paired samples. Statistical significance was
considered at the level of p<0.05 value. The SPSS-13 statistical
package was used for statistical analysis.
Results
JAK2 mutation status detection. Nine MPN patients out of eighteen were found positive for the JAK2V617F
point mutation (2 PV patients, 1 IMF patient and 6 ET patients) while
nine MPN patients (8 ET patients and 1 IMF patient), and the four ND
were JAK2V617F negative.
Apparently, two JAK2 WT patients were tested positive for the CALR
mutation and none positive for cMPL mutation. We then subjected the
samples that were positive for JAK2V617F
mutation to RFLP assays by using the BsaXI restriction enzyme, to
determine their ploidy status. All out of nine patients tested were
heterozygous for the JAK2V617F point mutation.
The presence of the JAK2V617F point mutation enhances in-vitro OCL forming ability. OCLs were formed from monocyte cells derived from ND, and JAK2 mutated or unmutated MPN patients (an example is given in Figure 1A). OCLs derived from heterozygous for the JAK2V617F mutation (Het) MPN patients, were significantly more compared to those from the JAK2 wild type (WT) MPN patients and ND as well; Median OCLs per 10.000 monocytes, (min- max): JAK2V617F (Het) MPN: 960 (159-3674), JAK2 WT MPN: 204 (81-1286) and p=0.03 Mann-Whitney test (Figure 1B). Considering ND the Median (min-max) OCLs per 10.000 monocytes were: 322 (215-522) and p=0.05 when compared to JAK2V617F (Het) MPN patients (Figure 1B).
OCLs
derived from JAK2V617F(Het) monocytes are enriched for the mutated
clone compared to the input monocyte cells reflecting an increased
outgrowth of the JAK2 V617F mutated clone. We then performed
quantitative Taqman based RT-PCR assays at genomic DNA level, in both
the input monocyte cells and in the OCLs that were finally derived from
them, aiming to clarify whether the increased OCL formation noted from JAK2V617F (Het) MPN monocytes reflects outgrowth of the wild type or the mutated clone. In all out of seven JAK2V617F (Het) MPN patients successfully tested, the ratio of JAK2V617F/JAK2WT genomic DNA was increased in the OCLs fraction compared to the input monocyte cells. The median enrichment of OCLs for the JAK2V617F mutated clone was 12% (range: 3.3%-26%) and p<0.01, Wilconxon Rank test (Figure 1C).
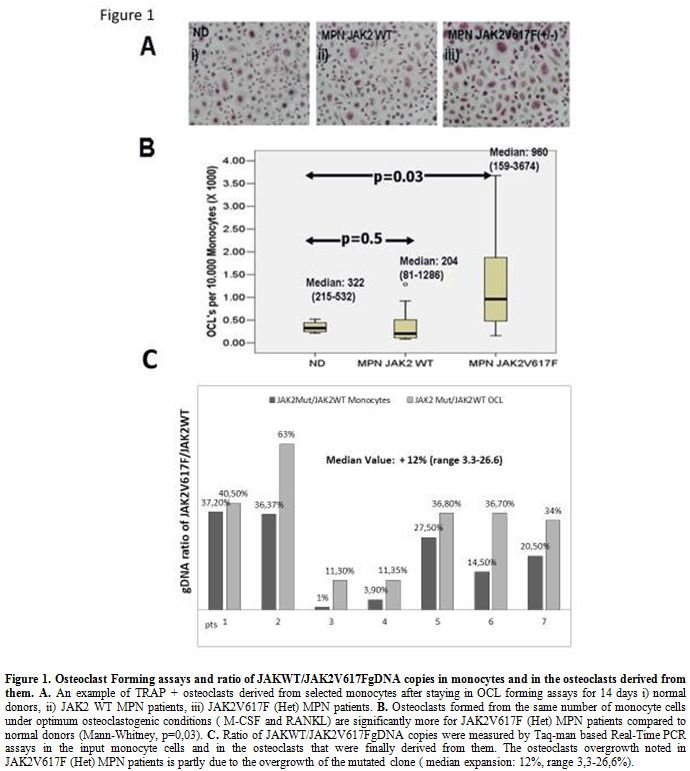 |
Figure 1. Osteoclast Forming assays and ratio of JAKWT/JAK2V617FgDNA copies in monocytes and in the osteoclasts derived from them. A.
An example of TRAP + osteoclasts derived from selected monocytes after
staying in OCL forming assays for 14 days i) normal donors, ii) JAK2 WT
MPN patients, iii) JAK2V617F (Het) MPN patients. B.
Osteoclasts formed from the same number of monocyte cells under optimum
osteoclastogenic conditions ( M-CSF and RANKL) are significantly more
for JAK2V617F (Het) MPN patients compared to normal donors
(Mann-Whitney, p=0,03). C.
Ratio of JAKWT/JAK2V617FgDNA copies were measured by Taq-man based
Real-Time PCR assays in the input monocyte cells and in the osteoclasts
that were finally derived from them. The osteoclasts overgrowth noted
in JAK2V617F (Het) MPN patients is partly due to the overgrowth of the
mutated clone ( median expansion: 12%, range 3,3-26,6%).
|
No evidence by CTXβ of increased OCL activity in JAK2 V617F patients.
The ability of JAK2 (Het) for higher OCL formation ex vivo poses the
question if these patients have in higher levels of osteoclast activity
in vivo. Therefore we quantified in serum from our MPN patients the
levels of CTXβ by ELISA but we cannot find any difference among groups
[median Value (min-
max) JAK2 (Het): 1,013(0,9-2,1 ng/ml) JAK2
WT: 1,014(0,78-2,1 ng/ml, p=0,69 Data not shown).
In Vitro Osteoclastogenesis from monocyte cells derived from MPN patients is dependent upon JAK2 function.
In order to address the dependence of osteoclastogenesis from JAK2
kinase in mutated and unmutated cells we performed OCL forming assays
under titrated inhibition of the JAK2 kinase by the selective JAK2
inhibitor AG-
490. We determined IC50 in OCL forming assays performed from 3 ND monocytes at 20μΜ (Figure 2A).
Afterwards, we performed OCL inhibition assays under titrated
concentrations of AG490 at 2μΜ and 20μΜ in MPN patients. OCL inhibition
is dose dependent in both groups of MPN patients (Table 1)
but it has a significantly suppressing effect on OCL formation only in
JAK2 (Het) MPN patients compared to ND. Of Note, at the higher
inhibitory concentration of 20 μΜ OCL formation in samples from
JAK2V617F (Het) patients was significantly reduced compared to ND (Table 1, p=0,02) and compared to JAK2 WT MPN patients as well (p=0,05). There was no statistically significant difference between ND and JAK2 WT MPN (p=0,28, Figure 2B).
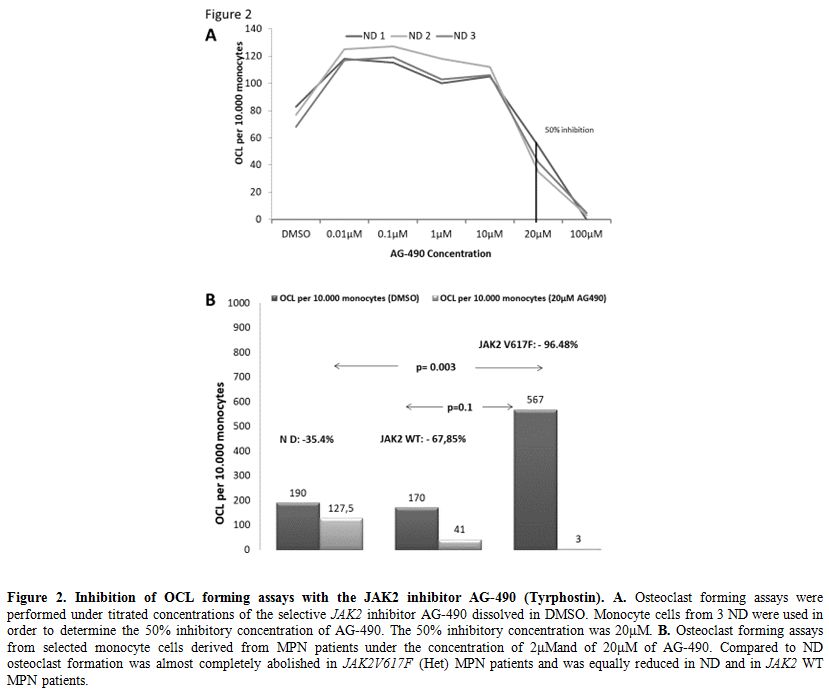 |
Figure 2.
Inhibition of OCL forming assays with the JAK2 inhibitor AG-490 (Tyrphostin). A. Osteoclast forming assays were performed under titrated concentrations of the selective JAK2
inhibitor AG-490 dissolved in DMSO. Monocyte cells from 3 ND were used
in order to determine the 50% inhibitory concentration of AG-490. The
50% inhibitory concentration was 20μΜ. B.
Osteoclast forming assays from selected monocyte cells derived from MPN
patients under the concentration of 2μΜand of 20μΜ of AG-490. Compared
to ND osteoclast formation was almost completely abolished in JAK2V617F (Het) MPN patients and was equally reduced in ND and in JAK2 WT MPN patients. |
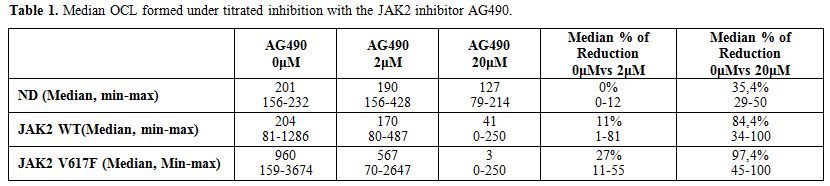 |
Table 1. Median OCL formed under titrated inhibition with the JAK2 inhibitor AG490. |
Discussion
In this study, we demonstrated that cells carrying the JAK2V617F point mutation are more efficient for osteoclast formation compared to cells carrying the JAK2 WT genotype. Monocytes derived from JAK2V617(Het) MPN patients can form 3 times more OCLs compared to the monocyte cells carrying the JAK2 WT genotype from both MPN patients and ND. Because input monocyte cells from JAK2V617(Het) patients is a mixture of cell clones carrying both JAK2V617F and JAK2 WT DNA
copies we performed real-time PCR at genome level to see whether the
OCL overgrowth is due to the preferential expansion of either of the
two clones. We demonstrated that the percentage of mutated JAK2
DNA copy numbers in the OCL fraction was increased compared to the
input monocyte cell fraction by a median value of 12%. In our culture
system the concentrations of RANKL and MCSF is optimum for monocyte
differentiation to osteoclasts, and therefore cells committed to the
OCL lineage are attached to the bottom of the plate, and they will
mature to OCL unless they do not survive. Hence, the enrichment of OCL
cell fraction for V617F mutated clones shows a proliferation/survival
advantage of the mutated clone compared to the JAK2 WT clones.
We
then tried to address if this ex vivo effect of JAK2V617F on OCL
formation has a detectable in vivo effect. Therefore we assessed
osteoclastogenic activity in the serum of our patients by measuring
CTXβ levels finding no difference between both groups. These data
suggest that the in vivo effect of JAK2V617F on OCL formation and
finally on the bones is not so robust in order to be detected by CTXβ
levels.
In order to provide further insight to the dependence of MPN osteoclastogenesis from the JAK2 kinase function, we performed OCL forming assays under titrated pharmacological inhibition of the JAK2 kinase with the selective JAK2 inhibitor AG-490 (Tyrphostin). We noted that osteoclastogenesis from cells carrying the JAK2V617F point mutation is more susceptible to JAK2
kinase inhibition than osteoclastogenesis from cells that are carrying
the wild type JAK2 kinase in a dose dependent manner. Our data provide
evidence for a therapeutic effect of JAK2 inhibitors in MPN patients
through reversion of OCL deregulation.
Our data enforce a model of
deregulated osteoclastogenesis in a disease characterised by the
genetically modified JAK2 kinase. In this study, we found out that the
acquisition of the JAK2V617F
point mutation in hematopoietic cells capable of osteoclastogenesis
increases their OCL forming ability, but these OCLs are also more
vulnerable to JAK2 kinase
inhibition. This poses to us the challenge to extrapolate the
underlying mechanism of increased osteoclastogenesis in JAK2V617F
mutated cells and furthermore to perform OCL functional assays in a
cohort of MPN patients enriched in patients homozygous for the JAK2
mutation. Targeting osteoclasts may be of value in MPN patients.
References
- Sanchez-Aguilera A, Mendez-Fer S. The hematopoietic
stem-cell niche in health and leukemia Cell. Mol. Life Sci. 2017,
74:579–590. https://doi.org/10.1007/s00018-016-2306-y PMid:27436341 PMCid:PMC5272896
- Mansour
A, Abou-Ezzi G, Sitnicka E, W Jacobsen SE, Wakkach A, Blin-Wakkach C.
Osteoclasts promote the formation of hematopoietic stem cell niches in
the bone marrow. J Exp Med. 2012 Mar 12;209(3):537-49. https://doi.org/10.1084/jem.20110994 PMid:22351931 PMCid:PMC3302238
- Lymperi
S, Ersek A, Ferraro F, Dazzi F, Horwood NJ. Inhibition of osteoclast
function reduces hematopoietic stem cell numbers in vivo. Blood. 2011
Feb 3;117(5):1540-9. https://doi.org/10.1182/blood- 2010-05-282855 PMid:21131587
- Miyamoto
K, Yoshida S, Kawasumi M, Hashimoto K, Kimura T, Sato Y, Kobayashi T,
Miyauchi Y, Hoshi H, Iwasaki R, Miyamoto H, Hao W, Morioka H, Chiba K,
Kobayashi T, Yasuda H, Penninger JM, Toyama Y, Suda T, Miyamoto T.
Osteoclasts are dispensable for hematopoietic stemcell maintenance and
mobilization. J Exp Med. 2011 Oct 24; 208(11):2175-81. https://doi.org/10.1084/jem.20101890 PMid:22006978 PMCid:PMC3201203
- Kollet
O, Dar A, Shivtiel S, Kalinkovich A, Lapid K, Sztainberg Y, Tesio M,
Samstein RM, Goichberg P, Spiegel A, Elson A, Lapidot T. Osteoclasts
degrade endosteal components and promote mobilization of hematopoietic
progenitor cells. Nat Med. 2006 Jun;12(6):657-64. https://doi.org/10.1038/nm1417 PMid:16715089
- Flores
C, Moscatelli I, Thudium CS, Gudmann NS, Thomsen JS, Bruel A, Karsdal
MA, Henriksen K, Richter J. Osteoclasts are not crucial for
hematopoietic stem cell maintenance in adult mice. Haematologica 2013,
98(12):1848–1855. https://doi.org/10.3324/haematol.2013.089466 PMid:24097632 PMCid:PMC3856959
- Rawlings JS, Rosler KM, Harrison DA, The JAK/STAT signaling pathway. J Cell Sci. 2004; 117: 1281– 1283. https://doi.org/10.1242/jcs.00963 PMID: 15020666
- Murray PJ, The JAK-STAT signaling pathway: input and output integration. J Immunol. 2007; 178: 2623–2629. https://doi.org/10.4049/jimmunol.178.5.2623 PMid:17312100
- Kwak
HB, Kim HS, Lee MS, Kim KJ, Choi EY, Choi MK, Kim JJ, Cho HJ, Kim JW,
Bae JM, Kim YK, Park BH, Ha H, Chun CH, Oh J. Pyridone6, a
pan-Janus-activated kinase inhibitor, suppresses
osteoclast
formation and bone resorption through down-regulation of receptor
activator of nuclear factor-kappaB (NF-kappaB) ligand (RANKL)-induced
c-Fos and nuclear factor of activated T cells (NFAT) c1 expression.
Biol Pharm Bull. 2009 Jan;32(1):45-50. https://doi.org/10.1248/bpb.32.45 PMid:19122279
- Li
CH, Zhao JX, Sun L, Yao ZQ, Deng XL, Liu R, Liu XY. AG490 inhibits
NFATc1expression and STAT3 activation during RANKL induced
osteoclastogenesis. Biochem Biophys Res Commun 2013 Jun 14:
435(4):533-9. https://doi.org/10.1016/j.bbrc.2013.04.084 PMid:23665018
- Jin-il
Park, Jennifer Lee, Mi-Ae Lim, Eun Kyung Kim, Sung Min Kim, JunGeolRyu,
Jae Ho Lee, Seung-Ki Kwok, kyung-Su Park, Ho- Youn Kim, Sung-Hwan Park,
Mi-La Cho. JAK2-STAT3 Blockade by AG490 Supresses autoimmune arthritis
in mice via reciprocal regulation of T cells and Th17 cells. Journal of
Immunology 2014; 192:4417-24. https://doi.org/10.4049/jimmunol.1300514 PMid:24688026
- Murakami
K, Kobayashi Y, Uehara S, Suzuki T, Koide M, Yamashita T, et al. A
Jak1/2 inhibitor, baricitinib, inhibits osteoclastogenesis by
suppressing RANKL expression in osteoblasts in vitro. PLoSONE 2017,
12(7): e0181126. https://doi.org/10.1371/journal. pone.0181126
- Mead
AJ, Chowdhury O, Pecquet C, Dusa A, Woll P, Atkinson D, Burns A, Score
J, Rugless M, Clifford R, Moule S, Bienz N, Vyas P, Cross N, Gale RE,
Henderson S, Constantinescu SN, Schuh A, Jacobsen SE. Impact of
germline JAKV617F mutation in human hematopoiesis. Blood. 2013 May
16;121(20):4156-65. https://doi.org/10.1182/blood-2012-05-430926 PMid:23535062
- Passamonti
F, Rumi E, Pietra D, Della Porta MG, Boveri E, Pascutto C, Vanelli L,
Arcaini L, Burcheri S, Malcovati L, Lazzarino M, Cazzola M. Relation
between JAK2 (V617F) mutation status, granulocyte activation, and
constitutive mobilization of CD34+ cells into peripheral blood in
myeloproliferative disorders. Blood. 2006 May 1;107(9):3676-82. https://doi.org/10.1182/blood-2005-09-3826 PMid:16373657
- Arranz
L, Sanchez-Aguilera A, Martin-Perez D, Isern J, Langa X, Tzankov A,
Lundberg P, Muntion S, Tzeng YS, Lai DM, Schwaller J, Skoda RC,
Mendez-Ferrer S (2014) Neuropathy of haematopoietic stem cell niche is
essential for myeloproliferative neoplasms. 512, Nature Aug 2014;
78–81. https://doi.org/10.1038/nature13383