This narrative review analyzes the relationship between gut microbiota and various immune-mediated inflammatory disorders in children, highlighting how microbial disruption may actively contribute to both disease phenotypes and therapeutic choices. Our search for pertinent studies in the principal biomedical databases included the terms "inflammasome", "familial Mediterranean fever", "juvenile idiopathic arthritis", "Kawasaki disease", and "IgA vasculitis”; studies dedicated to the pediatric age were included in our critical analysis. The results from our search highlighted significant points of contact between gut microbiota and pediatric immune-mediated inflammatory diseases, but - owing to inherent variability of the existing body of evidence - data results have been analyzed from a narrative standpoint, and not as a meta-analysis, for a constructive assessment of the role of intestinal microbial diversity and therapeutic modifications useful in different immune-mediated inflammatory disorders occurring in children. Additionally, this review examines the evolution of immune recognition mechanisms and their role in interactions between the host and the microbiota during childhood.
The Evolution of Immunologic Recognition Systems from Invertebrates to Humans
The recognition system that differentiates ‘self’ from ‘non-self’ has been integral to animal physiology since the emergence of multicellular organisms more than 600 million years ago.[3] Mechanisms involved in innate immunity differ from the adaptive immunity system that developed in the vertebrate lineage, and fundamentally serve as the initial basic line of defense against invaders to the human body.[4] Various receptors, ligands, and signaling molecules play a strategic role in the kick-off of many immune responses, triggering effector agents like interleukin (IL)-1, IL-6 and IL-12, which aim to the neutralization of pathogens and repair of injured tissues.[5] Interestingly, these primary defense tools have been identified in invertebrates, suggesting that different cellular products involved in immune reactions were highly preserved across species.[6] Indeed, genes that encode most innate immunity-related molecules share similarities with genes found in higher vertebrates, also indicating that elementary immune pathways may have evolved long before vertebrates appeared on the evolutionary timeline.[7] However, despite the presence of genes that impact both immunity and inflammatory response in invertebrates, the way they function may significantly differ from their counterparts in vertebrates, and it is likely that genes with a non-immunological role in invertebrates may have been repurposed for clearer immune functions in vertebrates, or vice versa, contributing to the complexity of long-term evolution.[8] These recognition systems have co-evolved with the human gut microbiota, ensuring balance and preventing pathologic dysbiosis.[9] Furthermore, the redefinition of initially non-immunologically active genes in invertebrates may have been a key-factor in the regulation of immunologic responses in higher vertebrates, shaping the dynamic interaction between hosts and their microbiota: this precondition upholds the role-playing game of immunity in maintaining health and stability of multicellular organisms.[10,11]Implications of Dysbiosis for the Progression of Immune-Mediated Disorders
The priming by which gut microbiota exerts its influence on innate immunity derives from disruption of the intestinal barrier integrity, directly involving the activity of immune cells and metabolite-mediated immune response modulation. To all intents and purposes, abnormalities in gut microbiota can activate Toll-like receptors (TLRs) and nucleotid-binding oligomerization domain proteins-like receptors (NLRs) by either increasing harmful metabolites or reducing beneficial products, such as short chain fatty acids (SCFA), ultimately leading to the overproduction of pro-inflammatory molecules like tumor necrosis factor (TNF)-α, IL-1β, and interferon (IFN)-γ.[12] Microbial communities have a typical stability under normal physiological conditions: however, they can undergo perturbations in response to Western dietary patterns, gastrointestinal infections, or administration of antibiotics.[13] An impaired function of the intestinal barrier triggers a pathophysiological cascade that affects various gastrointestinal and extra-intestinal disorders fueled by chronic inflammation through lymphatic and blood circulation.[14]Guittar and Shade observed consistent taxonomic and trait-based shifts in infant gut microbiota during the first 3 years of life: specifically, with respect to taxonomic composition, early succession was dominated by Bacteroidaceae and Bifidobacteriaceae, whereas the late succession was dominated by Lachnospiraceae, Ruminococcaceae, and again Bacteroidaceae in 56 infants from Finland and Estonia for whom longitudinal microbiome survey data were available.[15] Variations in gut microbiota may contribute to the development of peculiar autoimmune diseases: there are data related to type 1 diabetes, proving how the immune system mistakenly attacks insulin-producing beta cells of the pancreas following gut microbiota variations in genetically predisposed individuals, as a disrupted microbiota-immune interaction that affects Th1/Th17 balance may lead to production of autoreactive T lymphocytes and ultimately prime disease occurrence.[16] Specifically, intestinal dysbiosis in prediabetes may be characterized by reduced butyrate-producing bacteria such as Faecalibacterium prausnitzii and Roseburia, along with an increase of potentially harmful taxa such as Escherichia, Shigella, Shigella and Prevotella: this imbalance is associated with systemic inflammation and elevated levels of IL-6 and TNF-α. Moreover, an increased intestinal permeability might facilitate the translocation of bacterial components such as lipopolysaccharides, linking gut microbiota abnormalities to the development of insulin resistance and persistent hyperglycemia, as seen in type 2 diabetes.[17] Additional studies have also demonstrated that gut microbiota affects a wide range of clinical outcomes unrelated to intestinal disorders, and some have suggested that intestinal flora may influence many of severe neurological disorders through the brain-gut-microbe signaling system.[18] Moreover, early-life exposure to antibiotics can clearly disrupt gut microbiota, reduce microbial diversity, deplete key-commensal species, delay microbiota maturation, improperly train immune system development, weaken the intestinal barrier with subsequent potential inflammation, leading to adverse health effects in the long run.[19]
Exploring the Role of Gut Microbiota in Inflammasome-related Disorders
Twenty years ago the concept of autoinflammation was spread to define a group of complex conditions characterized by disrupted innate immunity and self-limited inflammatory bouts, involving skin, serosal membranes, joints, gut and other districts of the human body, which recur at a variable rhythmicity, sometimes concealed behind episodes of recurrent fevers in school-aged children.[20] The identification of causative genes directly involved in the innate immunity and specifically of pathogenic mutations within these genes coding for signaling complexes, receptors and sensors that are activated in response to pathogens and environmental factors led to a better understanding of how defense equipment may be regulated in normal conditions.[21] A prototypical hereditary autoinflammatory disorder is familial Mediterranean fever (FMF), caused by gain-of-function MEFV variants that affect pyrin function within the pyrin-inflammasome, characterized by mutant cells which become prone to uncontrolled pyroptotic death, clinically manifested by recurrent self-limiting attacks of fever and serositis.[22,23] In particular, Ozen et al. analyzed the gut microbiota of 28 FMF children from Turkey (plus 8 healthy controls) and 21 FMF patients from the United States of America (plus 6 controls) in 2021 and found neither significant differences between patients and controls, nor any differences in disease severity within each geographic area.[24] On the other hand, Delplanque et al observed a conceivable interplay between intestinal microbiota perturbations and phenotypic expression of FMF, analyzing a wider sample consisting of 119 FMF patients and 61 healthy controls: in particular, they found an over-representation of pro-inflammatory bacteria of the Enterobacteriaceae family in association with the most severe FMF phenotypes. In addition to direct immune modulation, alterations in the gut microbiota could trigger FMF attacks by impairing microbial metabolism and leading to the eventual expansion of pathogens. Moreover, it was speculated that FMF patients displaying the most severe clinical pictures, as well as colchicine resistance and/or homozygous MEFV pathogenic mutations, might display specific gut microbial signatures.[25] Unfortunately, these results have not been reproduced specifically for pediatric patients. Importantly, Pepoyan et al reported positive effects on C-reactive protein levels after administering the probiotic Lactobacillus acidophilus INMIA 9602 Er-2 strain 317/402 in 9 FMF patients (aged 18-50 years) living in Armenia; this was the first study to demonstrate the efficacy of a probiotic on disease activity in FMF patients.[26] The same authors found that the probiotic strain reduced the number of Candida albicans and Enterobacteriaceae in the gut of FMF patients[27] and that future placebo-controlled trials should help defining the usefulness of probiotic therapies, particularly focusing on Faecalibacterium, Blautia, and Clostridium species.[28] However, no data specifically referring to pediatric patients are currently available.Many inflammasome-related abnormalities with an aberrant pyroptosis pathway may have a bidirectional relationship with the gastrointestinal microflora, and an eventual dysbiosis could expose the host to pathogenic microorganisms and/or their metabolites, triggering inflammation. A specific group of rare hereditary autoinflammatory disorders characterized by recurrent inflammatory flares derives from the constitutive activation of the nucleotide-binding oligomerization domain protein-3 (NLRP3) inflammasome, driven by gain-of-function NLRP3 gene mutations and resulting in excessive IL-1β production.[29,30] The interplay between gut flora and host immune system may contribute to a kaleidoscopic clinical expression of different conditions in which the NLRP3 inflammasome is involved: for instance, Mafra et al explored this relationship in patients with chronic renal disease undergoing hemodialysis, who were found to have abundant Fusobacteria phyla - displaying a negative correlation with the mRNA of NLRP3 and IL-1β.[31] Identifying patients who may benefit from NLRP3 inflammasome-targeted therapies may be important in different clinical settings, though pediatric patients have never been tested. A very recent study by Deng et al. evaluated the effects of the enzymatically hydrolyzed small-molecule pectin derived from the leaves of Premna ligustroides Hemsl on the NLRP3 inflammasome, which is crucial to the initiation and progression of gout arthritis, also indicating that it could enhance uric acid metabolism.[32]
A rare but specific autoinflammatory disorder related to the NLR family caspase recruitment domain-containing protein 4 (NLRC4) inflammasome is caused by gain-of-function NLRC4 variants, leading to the constitutive activation of the NLRC4 inflammasome followed by early-onset enterocolitis and IL-18 oversecretion-based recurrent fevers, having the features of hemophagocytic lymphohistiocytosis.[33,34] However, experimental studies in NLRC4-mutated murine models have revealed that systemic high levels of IL-18 are gut microbiota-independent,[35] though a child with an NLRC4 mutation who was refractory to high-dose corticosteroids, cyclosporine, anakinra, and emapalumab required fecal microbiota transplant from an adult donor to manage a severely persistent diarrhea due to multidrug-resistant Enterobacter cloacae and Enterococcus faecalis; this treatment was associated with a definite control of inflammation and allowed tapering of the other concurrent immunosuppressive therapies.[36]
Loss of Microbial Diversity in the Pathogenic Loop of Juvenile Idiopathic Arthritis
Among trillions of bacteria present in different parts of the body, gut microbiota is the most studied in all rheumatologic diseases, and the link between gastrointestinal tube and immune system is further supported by the notion that disrupted populations of human gut bacteria can lead to immune-mediated processes involving both innate and adaptive immunity, as in the juvenile idiopathic arthritis (JIA) which is the most prevalent chronic arthritis affecting children below 16 years of age.[37] Indeed, evidence is increasing about changes of gut flora contributing to the development of JIA, making dysbiosis an occult driver of this immune-mediated condition. In particular, a dysregulated microbial metabolome would create a pro-inflammatory milieu, though no single intestinal organism has been identified in all performed studies as an unequivocal trigger of JIA.[38,39] In adults with rheumatoid arthritis, Faecalibacterium prausnitzii is consistently depleted, and that prevents its protective effect on the host, which is related to SCFA release, histone deacetylase inhibition, regulatory T (Treg) cells induction, secretion of microbial anti-inflammatory molecules, enhancement of epithelial barrier integrity, and modulation of pro- and anti-inflammatory cytokine responses.[40] Tejesvi et al. evaluated the fecal microbiota of 30 treatment-naive children with oligoarticular and polyarticular forms of JIA, finding lower abundance of Firmicutes and higher abundance of Bacteroidetes compared with controls; they also identified increased frequency of Actinobacteria and Fusobacteria and reduced frequency of Lentisphaerae, suggesting a distinct microbial profile for patients diagnosed with JIA.[41] In addition, Aggarwal et al. observed higher levels of Enterococcaceae species in the stools of children with enthesis-related JIA, whereas Prevotella was less abundant, apart from patients’ body mass indices and their HLA-B27 status.[42] In another study, van Dijkhuizen et al evaluated fecal samples from 78 treatment‐naive Italian and 21 treatment‐naive Dutch patients with non-systemic JIA, having less than 6 months of disease duration, comparing this double cohort to 107 geographically matched healthy children: importantly, they found Faecalibacterium prausnitzii increased in Italian patients at baseline, a species that could result in changes in SCFA production, while Allobaculum, Gemellaceae, Propionibacterium acnes, and Turicibacter were reduced compared to controls. Moreover, taxa such as Eggerthella, Rikenellaceae, Mogibacteriaceae, and Coprobacillus showed higher relative abundance in Dutch patients, though without reaching a statistical significance compared to controls.[43] Dong et al evaluated patients with systemic variant of JIA, nowadays named Still’s disease also in children, a condition primarily driven by IL-1, IL-6, and IL-18, characterized by systemic symptoms like fever and rash, which may progressively worsen until the development of macrophage activation syndrome and chronic arthritides:[44-46] The authors reported 17 patients with active disease, 15 with inactive disease, and 32 healthy controls, finding that Firmicutes and Bacteroidetes were the most abundant phyla in all patients’ samples and that the abundance of Bacteroidaceae was highest in the active Still group, followed by the inactive one.[47] Moreover, Gao et al investigated the hypothetical relationship between gut microbiota and JIA quantifying the potential role of plasma metabolites as disease mediators through the use of gut microbial genome-wide datasets encompassing 18,340 individuals from 24 cohorts, predominantly of European ancestry: two genera were positively associated with the risk of JIA: Rikenellaceae (OR = 1.199, 95% CI [1.034-1.190], p = 0.015) and Ruminococcus (OR = 1.401, 95% CI [1.024-1.916], p = 0.034), while Dorea showed a negative correlation with JIA; more specifically, metabolites associated with a decreased risk of JIA included cysteine-glutathione disulfide and dihydroxybenzoic acid, differently from furaneol sulfate which was associated to increased risk of JIA.[48] Little is known about the reciprocal influence between microbiota composition and immunosuppressive or biologic drugs used in JIA, and how this drug-microbiota interaction may have an impact on the clinical outcome. For instance, the effect of the most common anti-TNF-α drug used in children with JIA, etanercept, on the gut microbiota has not been extensively studied. However, Picchianti-Diamanti et al. found that etanercept could beneficially affect the microbiota and intestinal production of SCFAs in adults with rheumatoid arthritis, leading to increased distribution of Cyanobacteria, Nostocophycideae, and Nostocales, while Deltaproteobacteria and Clostridiaceae decreased.[49] This advantage may also be observed in patients with autoinflammatory disorders treated with etanercept, who may recover and partially restore their gut microbiota.[50-52] The plethora of environmental research undertaken in juvenile arthritides over the last decades, along with the challenges posed by using data from various patient cohorts, may enable microbiota-modifying strategies to restore a lost immune balance and improve the clinical response to canonical therapies.The Partnership of Gut Microbiota as a Driving Process in Vasculitic Syndromes
The gut microbiota is widely believed to be associated with immune-mediated vasculitides in childhood. An aberrant immune response involving genetic and environmental factors seems to play a seminal role in triggering Kawasaki disease (KD), a still mysterious vasculitis with a strong autoinflammatory component, which predominantly affects toddlers and specifically their medium-to-small size arteries: many attempts to develop universal scoring systems and detect KD children at higher risk of resistance to treatment have been unsuccessful,[53-55] and KD may present a different evolution according to yet unraveled demographic, genetic/epigenetic, or environmental factors like peculiar changes in the gut microbiota.[56-58]Differences in microbiota composition have been found in children with KD, in both acute and non-acute phases, with abnormal colonization of the intestinal tract by Streptococcus species and a wider presence of Gram-positive cocci in jejunal biopsies. In particular, a higher rate of Gram-positive cocci (of the genera Streptococcus and Staphylococcus), Eubacterium, Peptostreptococcus, and HSP60-producing Gram-negative microbes have been found in the feces of children with KD.[59] Yang et al found gut microbiota dysbiosis in children with KD during the acute phase, particularly a downregulation of SCFA-producing microbiota and over-proliferation of opportunistic pathogens.[60] Teramoto et al analyzed fecal DNA from 26 children with KD and 57 age-matched healthy controls, focusing on the microbial composition one year post-diagnosis. Even after clinical recovery, the KD group showed significant differences in microbial diversity, suggesting that the disease may have long-lasting effects on gut microbial balance. One of the main findings was a decrease in the abundance of Blautia, a genus known to produce butyrate, a SCFA with anti-inflammatory effects. Indeed, butyrate plays a role in maintaining intestinal epithelial integrity and promoting the development of Treg cells. This reduction of butyrate-producing bacteria suggests a loss of protective anti-inflammatory mechanisms in the gut, which may contribute to prime systemic inflammation. Conversely, there was a notable increase in Ruminococcus gnavus, a species associated with inflammatory bowel disease and rheumatoid arthritis: Ruminococcus gnavus is known to produce pro-inflammatory polysaccharides that can stimulate the innate immune system, particularly through TLR signaling pathways. Its abundance in KD patients supports the theory that a microbial trigger may amplify innate immune responses, thereby promoting vascular inflammation in the acute phase of KD. These findings confirm that gut dysbiosis may not only reflect the inflammatory milieu of KD, but actively participate in initiating or sustaining KD immune dysregulation, especially through the loss of anti-inflammatory taxa and rise of pro-inflammatory bacteria, highlighting the idea that targeting gut microbiota - through probiotics and prebiotics - may represent an adjunctive strategy in the management of children with KD.[61]
Further studies have confirmed that an imbalance in gut microbiota might indirectly interfere with a regular function of innate and adaptive immunity, and that variable microbiota interactions with environmental factors, mainly infectious agents, can selectively steer the development of Behçet’s disease (BD), another systemic vasculitis of unknown etiology characterized by mucocutaneous manifestations, including recurrent oral and genital ulcerations, ocular manifestations, especially chronic relapsing uveitis, and systemic involvement of arteries and veins of all sizes, which might have a pediatric onset: BD has several autoinflammatory features, including recurrent self-limited clinical manifestations overlapping with monogenic autoinflammatory disorders due to abnormally increased inflammatory responses with a robust innate component.[62] The gut microbiota of BD patients lacks butyrate-producing bacteria and methanogenic bacteria: Consolandi et al first reported the microbiota characteristics in BD, showing a significant depletion of the genera Roseburia and Subdoligranulum and also significantly reduced levels of butyrate production.[63] Unfortunately, there are no studies conducted specifically in children with BD.
Immunoglobulin A vasculitis (IgAV) is the most common primary vasculitis occurring in children, triggered by a series of abnormal immune responses primed by infections, drugs, vaccines, parasites or even tumors in genetically predisposed individuals: the disease typically follows a self-limiting course, with symptoms resolving within a few weeks to months, but one third of pediatric patients with IgAV may have a recurrence of the vasculitis picture.[64] There is increasing evidence suggesting that bacterial dysbiosis with dominant Bacteroidaceae and decreased Coriobacteriaceae may influence the progression and the recurrence rates of IgAV.[65] Wang et al studied the composition of gut microbiota by 16S rRNA gene-based sequencing in 85 children with IgAV, finding abundant Fusobacteria and also a decrease of Firmicutes.[66] Further studies have demonstrated Th1/Th2 and Treg/Th17 imbalances in IgAV, as evidenced by the suppression of Th1 and Treg cells and upregulated Th2 and Th17 differentiation: in particular, Liang M et al studied blood and stool specimens of 27 children with IgAV, showing higher serum levels of IFN-γ, IL-4, IL-10, and IL-17. The authors correlated the prevalence of Blautia and specifically of Blautia wexlerae to serum IL-10 levels.[67] Additionally, Liang W et al examined gut microbiota composition of 127 children with IgAV, finding the reduction of four genera, namely Lachnospira, Ruminococcus, Roseburia, and Streptococcus, potentially implicated in the production of the anti-inflammatory SCFAs butyrate and propionate.[68] Table 1 shows a list of some pediatric non-intestinal immune-mediated inflammatory disorders influenced by microbiota changes and subsequent immunological effects or clinical implications. Figure 1 shows the mechanisms involved in the symbiotic relationship between gut microbiota and the immunologic axis, which helps maintaining homeostasis in the host: an eventual dysbiosis (starting in the pediatric age) with barrier disruption, decreased production of SCFA, and increased release of pro-inflammatory cytokines may induce specific disease patterns, as found in FMF, JIA, KD, and IgAV.
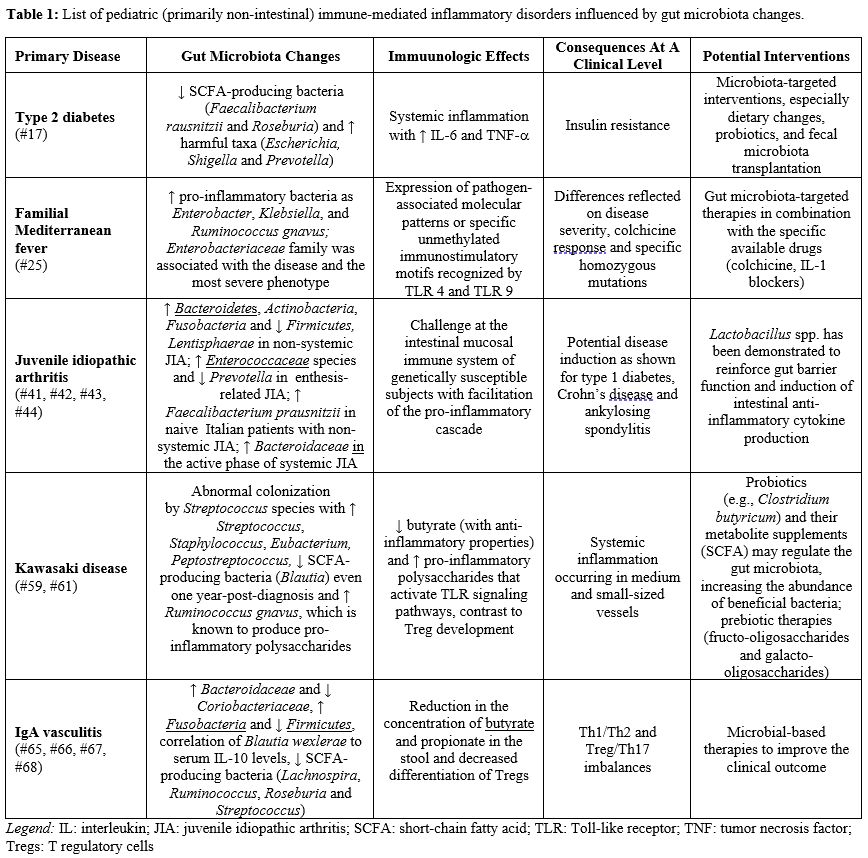 |
Table 1. List of pediatric (primarily non-intestinal) immune-mediated inflammatory disorders influenced by gut microbiota changes. |
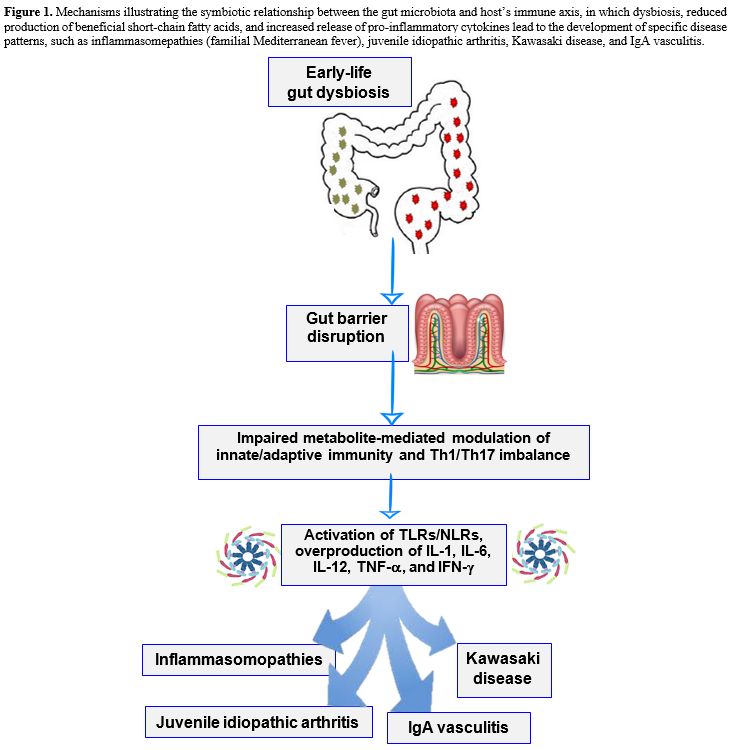 |
Figure 1. Mechanisms illustrating the symbiotic relationship between the gut microbiota and host’s immune axis, in which dysbiosis, reduced production of beneficial short-chain fatty acids, and increased release of pro-inflammatory cytokines lead to the development of specific disease patterns, such as inflammasomepathies (familial Mediterranean fever), juvenile idiopathic arthritis, Kawasaki disease, and IgA vasculitis. |
Probiotics as Training Tools to Prime the Progress of Innate Immunity of Children
An early administration of Gram-positive bacteria, including species from the Lactobacillus and Bifidobacterium genera, as well as Enterococci or yeast species such as Saccharomyces boulardii, collectively called ‘probiotics,’ helps restoring physiologic microbial balance, supports innate immunity in young patients, and promotes immunological tolerance to prevent undesirable immune responses against commensal bacteria whose cellular components may share certain features with pathogenic ones.[69] A recent randomized controlled trial strongly supported this therapeutic approach, demonstrating that a probiotic mixture containing Bifidobacterium breve, Bifidobacterium lactis, and Lactobacillus rhamnosus can give benefits to children with history of upper respiratory tract infections.[70] The study also confirmed previous results, highlighting the importance of probiotic competitive adherence to the intestinal mucosa to confer an advantage to the host.[71] Similarly, a Cochrane systematic review showed that probiotics can lower the incidence and overall duration of upper respiratory tract infections, reduce antibiotic use, and decrease the number of school absence days, though further trials are needed to consolidate these promising findings. Additional mechanisms by which probiotics exert their benefits include modulation of innate immunity pathways, stimulating the release of antimicrobial proteins via interaction with pathogen sensors, such as TLRs and NLRs, and through a complex network of cytokines produced by innate immune cells, including IL-17 and IL-22, which are emerging as crucial regulators of antimicrobial-peptide production in the gut.[72] Future large trials should focus on identifying the optimal probiotic strains, dosing regimens, and durations of administration to maximize preventive or therapeutic outcomes across different pediatric scenarios.Conclusion and Future Directions
All living organisms can be regarded as complex entities colonized by diverse microbial ecosystems across nearly all body tissues (with the exception of the central nervous system). The gut is the most densely populated, with an estimated 100 trillion resident microorganisms in humans. However, the interdependence of different microbial communities in the gut flora remains unclear, and various approaches are being investigated, including the use of probiotics to supplement beneficial commensals, dietary modifications, and fecal microbial transplants.Indeed, a wealth of data supports the notion that disruptions in the diversity of human gut bacteria can have enduring effects on protean immunologic functions, particularly in the early developmental phase. Limitations of our present study include the paucity of pediatric cohorts reported in the published literature, small sample sizes, age differences among patients, and heterogeneity in microbiota assessment techniques. As investigations continue to reveal the intricate processes underlying microbiota-immunity interactions, targeted microbiota-modifying therapies could offer promising options for many complex diseases and improve patient outcomes. It is important to acknowledge that all studies conducted to date have been location-specific, whereas studies across continents should be prioritized to elucidate how the gut microbiota can cause specific organ inflammatory responses. Understanding the relationship between the gut microbiota and innate immunity may offer important insights into disease prevention and pave the way for more tailored treatment approaches.
Abbreviations
BD: Behçet’s disease; FMF: familial Mediterranean fever; IFN: interferon; IgAV: immunoglobulin A vasculitis; IL: interleukin; JIA: juvenile idiopathic arthritis; KD: Kawasaki disease; NLR: nucleotide-binding oligomerization domain protein-3; NLRP3: nucleotide-binding oligomerization domain protein-3; NLRC4: NLR family caspase recruitment domain-containing protein 4; SCFA: short-chain fatty acid; TLR: Toll-like receptor; TNF: tumor necrosis factor; Treg: T regulatory (cells)Author Contributions
All authors made substantial contributions to the conception and design of the review, the acquisition, analysis, and interpretation of the literature, as well as to the drafting and critical revision of the manuscript. All authors approved the final version of the manuscript and agree to be accountable for all aspects of the work, ensuring its accuracy and integrity.References
- Cai J, Sun L, Gonzalez F. Gut
microbiota-derived bile acids in intestinal immunity, inflammation, and
tumorigenesis. Cell Host Microbe 2022;3:289-300. https://doi.org/10.1016/j.chom.2022.02.004 PMid: 35271802
- Wu
H, Tremaroli V, Schmidt C, et al. Bäckhed, F. The gut microbiota in
prediabetes and diabetes: a population-based cross-sectional study.
Cell Metab 2020;32:379-90.e3. https://doi.org/10.1016/j.cmet.2020.06.011 PMid: 32652044
- Dzik JM. The ancestry and cumulative evolution of immune reactions. Acta Biochim Pol 2010;57:443-66. PMid: 21046016
- Janeway CA, Medzhitov R. Innate immune recognition. Annu Rev Immunol 2002;20:197-216. https://doi.org/10.1146/annurev.immunol.20.083001.084359 PMid: 11861602
- Li D, Wu M. Pattern recognition receptors in health and diseases. Signal Transuct Target Ther 2021;6:291. https://doi.org/10.1038/s41392-021-00687-0 PMid: 34344870
- Hoffmann JA, Reichhart JM. Drosophila innate immunity: an evolutionary perspective. Nat Immunol 2002;3:121-6. https://doi.org/10.1038/ni0202-121 PMid: 11812988
- Litman GW, Cannon JP, Dishaw LJ. Reconstructing immune phylogeny: new perspectives. Nat Rev Immunol 2005;5:866-79. https://doi.org/10.1038/nri1712 PMid: 16261174
- Pancer Z, Cooper MD. The evolution of adaptive immunity. Annu Rev Immunol 2006;24:497-518. https://doi.org/10.1146/annurev.immunol.24.021605.090542 PMid: 16551257
- Hand
TW, Vujkovic-Cvijin I, Ridaura VK, et al. Linking the microbiota,
chronic disease, and the immune system. Trends Endocrinol Metab
2016;27:831-43. https://doi.org/10.1016/j.tem.2016.08.003 PMid: 27623245
- Belkaid Y, Hand TW. Role of the microbiota in immunity and inflammation. Cell 2014;157:121-31. https://doi.org/10.1016/j.cell.2014.03.011 PMid: 24679531
- McFall-Ngai M. Adaptive immunity: care for the community. Nature 2007;445(7124):153. https://doi.org/10.1038/445153a PMid: 17215830
- Capaldo
CT, Beeman N, Hilgarth RS, et al. IFN-γ and TNF-α-induced GBP-1
inhibits epithelial cell proliferation through suppression of
β-catenin/TCF signaling. Mucosal Immunol 2012;5:681-90. https://doi.org/10.1038/mi.2012.41 PMId: 22692453
- Gueddouri
D, Caüzac M, Fauveau V, et al. Insulin resistance per se drives early
and reversible dysbiosis-mediated gut barrier impairment and
bactericidal dysfunction. Mol Metab 2022;57:101438. https://doi.org/10.1016/j.molmet.2022.101438 PMid: 35007789
- Camilleri M. Leaky gut: mechanisms, measurement and clinical implications in humans. Gut 2019;68:1516-26. https://doi.org/10.1136/gutjnl-2019-318427 PMid: 31076401
- Guittar J, Shade A. Trait-based community assembly and succession of the infant gut microbiome. Nat Commun 2019;10:512. https://doi.org/10.1038/s41467-019-08377-w PMid: 30710083
- Faith JJ, Guruge JL, Charbonneau M, et al. The long-term stability of the human gut microbiota. Science 2013;341:1237439. https://doi.org/10.1126/science.1237439 PMid: 23828941
- Liberty
IA, Hanifah L, Rachmat HF, et al. Implication of taxonomic abundance of
gut microbiota in prediabetes: a systematic review. Front Nutr
2025;12:1577528. https://doi.org/10.3389/fnut.2025.1577528 PMid: 40308637
- He
M, Liu L, Li Y, et al. Formation of gut microbiota and application of
microecological regulators in children. Transl Pediatr 2025;14:2023-41.
https://doi.org/10.21037/tp-2025-303 PMid: 40949907
- Zhou
W, Sailani MR, Contrepois K, et al. Longitudinal multi-omics of
host-microbe dynamics in prediabetes. Nature 2019;569:663-71. https://doi.org/10.1038/s41586-019-1236-x PMid: 31142858
- Cantarini
L, Iacoponi F, Lucherini OM, et al. Validation of a diagnostic score
for the diagnosis of autoinflammatory diseases in adults. Int J
Immunopathol Pharmacol 2011;24:695-702. https://doi.org/10.1177/039463201102400315 PMid: 21978701
- Sarı İ, Birlik M, Kasifoğlu T. Familial Mediterranean fever: an updated review. Eur J Rheumatol 2014;1:21-33. https://doi.org/10.5152/eurjrheum.2014.006 PMid: 27708867
- Rigante
D, Frediani B, Galeazzi M, et al. From the Mediterranean to the sea of
Japan: the transcontinental odyssey of autoinflammatory diseases.
Biomed Res Int 2013;2013:485103. https://doi.org/10.1155/2013/485103 PMid: 23971037
- Kisla Ekinci RM, Kilic Konte E, Akay N, et al. Familial Mediterranean fever in childhood. Turk Arch Pediatr 2024;59:527-34. https://doi.org/10.5152/TurkArchPediatr.2024.24188 PMid: 39540697
- Ozen
S, Lutz HL, Rivera VM, et al. Microbiome is not linked to clinical
disease severity of familial Mediterranean fever in an international
cohort of children. Clin Exp Rheumatol 2021;39 Suppl 132(5):102-8. https://doi.org/10.55563/clinexprheumatol/olvbyd PMid: 34251310
- Delplanque
M, Benech N, Rolhion N, et al. Gut microbiota alterations are
associated with phenotype and genotype in familial Mediterranean fever.
Rheumatology (Oxford) 2024;63:1039-48. https://doi.org/10.1093/rheumatology/kead322 PMid: 37402619
- Pepoyan
AZ, Balayan MH, Manvelyan AM, et al. Lactobacillus acidophilus INMIA
9602 Er-2 strain 317/402 probiotic regulates growth of commensal
Escherichia coli in gut microbiota of familial Mediterranean fever
disease subjects. Lett Appl Microbiol 2017;64:254-60. https://doi.org/10.1111/lam.12722 PMid: 28140472
- Pepoyan
A, Balayan M, Manvelyan A, et al. Probiotic Lactobacillus acidophilus
strain INMIA 9602 Er 317/402 administration reduces the numbers of
Candida albicans and abundance of enterobacteria in the gut microbiota
of familial Mediterranean fever patients. Front Immunol 2018;9:1426. https://doi.org/10.3389/fimmu.2018.01426 PMid: 29997616
- Pepoyan
E, Marotta F, Manvelyan A, et al. Placebo-resistant gut bacteria:
Akkermansia muciniphila spp. and familial Mediterranean fever disease.
Front Cell Infect Microbiol 2024;14:1336752. https://doi.org/10.3389/fcimb.2024.1336752 PMid: 38465231
- Rigante
D. The protean visage of systemic autoinflammatory syndromes: a
challenge for inter-professional collaboration. Eur Rev Med Pharmacol
Sci 2010;14:1-18. PMid: 20184084
- Yüksel
Ş, Eren E, Hatemi G, et al. Novel NLRP3/cryopyrin mutations and
pro-inflammatory cytokine profiles in Behçet's syndrome patients. Int
Immunol 2014;26:71-81. https://doi.org/10.1093/intimm/dxt046 PMid: 24135410
- Mafra
D, Alvarenga L, Cardozo LFMF, et al. Gut microbiota and NLRP3
inflammasome activation in hemodialysis patients: exploring the link
with systemic inflammation. Mol Biol Rep 2025;52:465. https://doi.org/10.1007/s11033-025-10562-8 PMid: 40387945
- Deng
L, He R, Mao J, et al. Modulation of NLRP3 inflammasome and uric acid
metabolism by small molecule pectin from Premna ligustroides Hemsl
leaves: implications for hyperuricemia management. Int J Biol Macromol
2025;331(Pt 1):148214. https://doi.org/10.1016/j.ijbiomac.2025.14821 PMid: 41075904
- Rigante
D. A systematic approach to autoinflammatory syndromes: a spelling
booklet for the beginner. Expert Rev Clin Immunol 2017;13:571-97. https://doi.org/10.1080/1744666X.2017.1280396 PMid: 28064547
- Stabile
A, Bertoni B, Ansuini V, et al. The clinical spectrum and treatment
options of macrophage activation syndrome in the pediatric age. Eur Rev
Med Pharmacol Sci 2006;10:53-9. PMid: 16705949
- Eeckhout E, Asaoka T, Van Gorp H, et al. The autoinflammation-associated NLRC4V341A
mutation increases microbiota-independent IL-18 production but does not
recapitulate human autoinflammatory symptoms in mice. Front Immunol
2023;14:1272639. https://doi.org/10.3389/fimmu.2023.1272639 PMid: 38090573
- Bracaglia
C, Marucci G, Del Chierico F, et al. Microbiota transplant to control
inflammation in a patient with NLRC4 gain-of-function–induced disease.
J Allergy Clin Immunol 2023;152:302-3. https://doi.org/10.1016/j.jaci.2023.03.031 PMid: 37178069
- Verwoerd
A, ter Haar NM, de Roock S, et al. The human microbiome and juvenile
idiopathic arthritis. Pediatr Rheumatol Online J 2016;14:55. https://doi.org/10.1186/s12969-016-0114-4 PMid: 27650128
- Rigante D. The fresco of autoinflammatory diseases from the pediatric perspective. Autoimmun Rev 2012;11:348-56. https://doi.org/10.1016/j.autrev.2011.10.008 PMid: 22024500
- Shibahara
T, Temizoz B, Egashira S, et al. Microbial dysbiosis fuels STING-driven
autoinflammation through cyclic dinucleotides. J Autoimmun
2025;154:103434. https://doi.org/10.1016/j.jaut.2025.103434 PMid: 40334619
- Han
EJ, Ahn JS, Chae YJ, et al. Immunomodulatory roles of Faecalibacterium
prausnitzii and Akkermansia muciniphila in autoimmune diseases:
mechanistic insights and therapeutic potential. Clin Rev Allergy
Immunol 2025;68:77. https://doi.org/10.1007/s12016-025-09093-8 PMid: 40759811
- Tejesvi
MV, Arvonen M, Kangas SM, et al. Faecal microbiome in new‐onset
juvenile idiopathic arthritis. Eur J Clin Microbiol Infect Dis
2016;35:363-70. https://doi.org/10.1007/s10096-015-2548-x PMid: 26718942
- Aggarwal
A, Sarangi AN, Gaur P, et al. Gut microbiome in children with
enthesitis-related arthritis in a developing country and the effect of
probiotic administration. Clin Exp Immunol 2017;187:480-9. https://doi.org/10.1111/cei.12900 PMid: 27861762
- van
Dijkhuizen EHP, Del Chierico F, Malattia C, et al. Microbiome analytics
of the gut microbiota in patients with juvenile idiopathic arthritis: a
longitudinal observational cohort study. Arthritis Rheumatol
2019;71:1000-10. https://doi.org/10.1002/art.40827 PMid: 30592383
- Rigante
D. Autoinflammatory syndromes behind the scenes of recurrent fevers in
children. Med Sci Monit 2009;15:RA179-87. PMid: 19644432
- Rogowska
J, Kubicka J, Grabowska M, et al. Characterization of patients with
macrophage activation syndrome secondary to systemic juvenile
idiopathic arthritis. Clin Rheumatol 2025;44:2023-9. https://doi.org/10.1007/s10067-025-07420-0 PMid: 40146446
- Rigante
D. The broad-ranging panorama of systemic autoinflammatory disorders
with specific focus on acute painful symptoms and hematologic
manifestations in children. Mediterr J Hematol Infect Dis
2018;10(1):e2018067. https://doi.org/10.4084/MJHID.2018.067 PMid: 30416699
- Dong
YQ, Wang W, Li J, et al. Characterization of microbiota in
systemic-onset juvenile idiopathic arthritis with different disease
severities. World J Clin Cases 2019;7:2734-45. https://doi.org/10.12998/wjcc.v7.i18.2734 PMid: 31616689
- Gao
B, Wang Z, Wang K, et al. Relationships among gut microbiota, plasma
metabolites, and juvenile idiopathic arthritis: a mediation Mendelian
randomization study. Front Microbiol 2024;15:1363776. https://doi.org/10.3389/fmicb.2024.1363776 PMid: 38605717
- Picchianti-Diamanti
A, Panebianco C, Salemi S, et al. Analysis of gut microbiota in
rheumatoid arthritis patients: disease-related dysbiosis and
modifications induced by etanercept. Int J Mol Sci 2018;19:2938. https://doi.org/10.3390/ijms19102938 PMid: 30261687
- Federico
G, Rigante D, Pugliese AL, et al. Etanercept induces improvement of
arthropathy in chronic infantile neurological cutaneous articular
(CINCA) syndrome. Scand J Rheumatol 2003;32:312-4. https://doi.org/10.1080/03009740310003974 PMid: 14690147
- Kusuhara
K, Hoshina T, Saito M, et al. Successful treatment of a patient with
tumor necrosis factor receptor-associated periodic syndrome using a
half-dose of etanercept. Pediatr Int 2012;54:552-5. https://doi.org/10.1111/j.1442-200X.2011.03525.x PMid: 22830546
- Cantarini
L, Rigante D, Lucherini OM, et al. Role of etanercept in the treatment
of tumor necrosis factor receptor-associated periodic syndrome:
personal experience and review of the literature. Int J Immunopathol
Pharmacol 2010;23:701-7. https://doi.org/10.1177/039463201002300303 PMid: 20943039
- De
Rosa G, Pardeo M, Rigante D. Current recommendations for the
pharmacologic therapy in Kawasaki syndrome and management of its
cardiovascular complications. Eur Rev Med Pharmacol Sci 2007;11:301-8.
PMid: 18074939
- Türkuçar
S, Yildız K, Acari C, et al. Risk factors of intravenous immunoglobulin
resistance and coronary arterial lesions in Turkish children with
Kawasaki disease. Turk J Pediatr 2020;62:1-9. https://doi.org/10.24953/turkjped.2020.01.001 PMid: 32253860
- Rigante
D, Valentini P, Rizzo D, et al. Responsiveness to intravenous
immunoglobulins and occurrence of coronary artery abnormalities in a
single-center cohort of Italian patients with Kawasaki syndrome.
Rheumatol Int 2010;30:841-6. https://doi.org/10.1007/s00296-009-1337-1 PMid: 20049445
- Rigante
D. Kawasaki disease as the immune-mediated echo of a viral infection.
Mediterr J Hematol Infect Dis 2020;12(1):e2020039. https://doi.org/10.4084/MJHID.2020.039 PMid: 32670517
- Rigante
D, Tarantino G, Valentini P. Non-infectious makers of Kawasaki
syndrome: tangible or elusive triggers? Immunol Res 2016;64:51-4. https://doi.org/10.1007/s12026-015-8679-4 PMid: 26232895
- Uysal
F, Bostan OM, Celebi S, et al. Outcomes of Kawasaki disease: a
single-center experience. Clin Pediatr (Phila) 2015;54:579-84. https://doi.org/10.1177/0009922814561594 PMid: 25475592
- Esposito
S, Polinori I, Rigante D. The gut microbiota-host partnership as a
potential driver of Kawasaki syndrome. Front Pediatr 2019;7:124. https://doi.org/10.3389/fped.2019.00124 PMid: 31024869
- Yang
Q, Kang Y, Tang W, et al. Interplay of gut microbiota in Kawasaki
disease: role of gut microbiota and potential treatment strategies.
Future Microbiol 2025;20:357-69. https://doi.org/10.1080/17460913.2025.2469432 PMid: 40013895
- Teramoto
Y, Akagawa S, Hori SI, et al. Dysbiosis of the gut microbiota as a
susceptibility factor for Kawasaki disease. Front Immunol
2023;14:1268453. https://doi.org/10.3389/fimmu.2023.1268453 PMid: 38022552
- Gül A. Pathogenesis of Behçet's disease: autoinflammatory features and beyond. Semin Immunopathol 2015;37:413-8. https://doi.org/10.1007/s00281-015-0502-8 PMId: 26068404
- Consolandi
C, Turroni S, Emmi G, et al. Behçet's syndrome patients exhibit
specific microbiome signature. Autoimmun Rev 2015; 14:269-76. https://doi.org/10.1016/j.autrev.2014.11.009 PMid: 25435420
- Chen
JY, Mao JH. Henoch-Schönlein purpura nephritis in children: incidence,
pathogenesis and management. World J Pediatr 2015;11:29-34. https://doi.org/10.1007/s12519-014-0534-5 PMid: 25557596
- Wen
M, Dang X, Feng S, et al. Integrated analyses of gut microbiome and
host metabolome in children with Henoch-Schönlein purpura. Front Cell
Infect Microbiol 2022;11:796410. https://doi.org/10.3389/fcimb.2021.796410 PMid: 35145922
- Wang
X, Zhang L, Wang Y, et al. Gut microbiota dysbiosis is associated with
Henoch-Schönlein purpura in children. Int Immunopharmacol 2018; 58:1-8.
https://doi.org/10.1016/j.intimp.2018.03.003 PMId: 29525681
- Liang
M, Deng Z, Wu W, et al. Study on the correlation between intestinal
flora and cytokines in children with Henoch-Schönlein purpura. Cytokine
2025;191:156959. https://doi.org/10.1016/j.cyto.2025.156959 PMid: 40373421
- Liang
Y, Zhao C, Zhao L, et al. Taxonomic and functional shifts of gut
microbiome in immunoglobulin A vasculitis children and their mothers.
Front Pediatr 2024;12:1356529. https://doi.org/10.3389/fped.2024.1356529 PMid: 38410769
- Mazziotta
C, Tognon M, Martini F, et al. Probiotics mechanism of action on immune
cells and beneficial effects on human health. Cells 2023;12:184. https://doi.org/10.3390/cells12010184 PMid: 36611977
- Bettocchi
S, Comotti A, Elli M, et al. Probiotics and fever duration in children
with upper respiratory tract infections: a randomized clinical trial.
JAMA Netw Open 2025;8:e2500669. https://doi.org/10.1001/jamanetworkopen.2025.0669 PMid: 40085083
- Bermudez-Brito M, Plaza-Díaz J, Muñoz-Quezada S, et al. Probiotic mechanisms of action. Ann Nutr Metab 2012;61:160-74. https://doi.org/10.1159/000342079 PMid: 23037511
- Hao
Q, Dong BR, Wu T. Probiotics for preventing acute upper respiratory
tract infections. Cochrane Database Syst Rev 2022;8:CD006895. https://doi.org/10.1002/14651858.CD006895.pub4 PMid: 32670517